
Резекция носовой перегородки, как исправляют искривление перегородки?
Искривление носовой перегородки в разной степени выраженности отмечается почти у 80% людей. Иногда эта анатомическая особенность служит причиной нарушения носового дыхания и способствует развитию других ЛОР-заболеваний. Такие ситуации являются показанием для проведения операции на перегородке носа.
Характер вмешательства зависит от типа и выраженности деформации внутриносовых структур. Самым часто используемым вариантом операции является резекция носовой перегородки.
Искривление перегородки носа бывает врожденным и даже наследственно обусловленным состоянием. При этом также могут отмечаться пороки развития других костей лицевого черепа. Но встречаются случаи приобретенного искривления перегородки.
Основные причины такой патологии
- неравномерный рост кости и хряща в пубертатном периоде, в результате чего быстро увеличивающаяся хрящевая часть перегородки начинает изгибаться;
- травмы в детском и подростковом возрасте, приводящие к нарушению функционирования ростковых зон;
- механическое воздействие избыточно разросшейся ткани при полипозе, аденоидите, гипертрофическом рините и риносинусите;
- гипертрофия сошниково-носового обонятельного органа Якобсона, который для человека является рудиментом и обычно остается недоразвитым.


Многие люди не знают об имеющейся у них асимметричности носовых ходов. Такое возможно, если искривленная носовая перегородка грубо не нарушает циркуляцию воздуха и отток из околоносовых пазух. Это состояние считается индивидуальной анатомической особенностью и не требует лечения.
Но в ряде случаев отклоненная от средней линии перегородка становится причиной предрасположенности к частым и рецидивирующим синуситам, отитам, хроническому риниту и другим заболеваниям ЛОР-органов. И самой частой жалобой пациентов становится затруднение носового дыхания. А при выраженных искривлениях могут отмечаться храп и внешние лицевые деформации. Все это является показанием для решения вопроса о резекции перегородки носа или других видах оперативного вмешательстваКаким бывает искривление
Искривлением называют отклонение (девиацию) носовой перегородки или ее части от средней линии.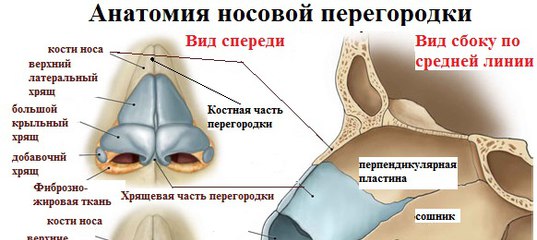 При этом деформация может иметь С-образный вид и напоминать дугу или же образовывать подобие буквы S. В последнем случае будет нарушена вентиляция обоих носовых ходов. Чаще всего девиации подвержена четырехугольная хрящевая часть перегородки, то есть ее передние 2/3.
При этом деформация может иметь С-образный вид и напоминать дугу или же образовывать подобие буквы S. В последнем случае будет нарушена вентиляция обоих носовых ходов. Чаще всего девиации подвержена четырехугольная хрящевая часть перегородки, то есть ее передние 2/3.
Про искривление говорят и в случае появления костных деформаций в виде гребней и шипов различной формы, положения и размера. Они образуются из костной части перегородки, чаще всего из края сошника. Эти образования могут вдаваться в слизистую оболочку или формировать дополнительные выступы внутри полости носа.
Посттравматические искривления обычно достаточно грубые, с резкими изломами и смещениями тканей. При этом нередко отмечается вывих четырехугольного хряща и нарушение его целостности.
Существует 3 степени выраженности искривления перегородки носа. Первая степень обычно не сопровождается явными клиническими признаками и характеризуется как незначительная деформация.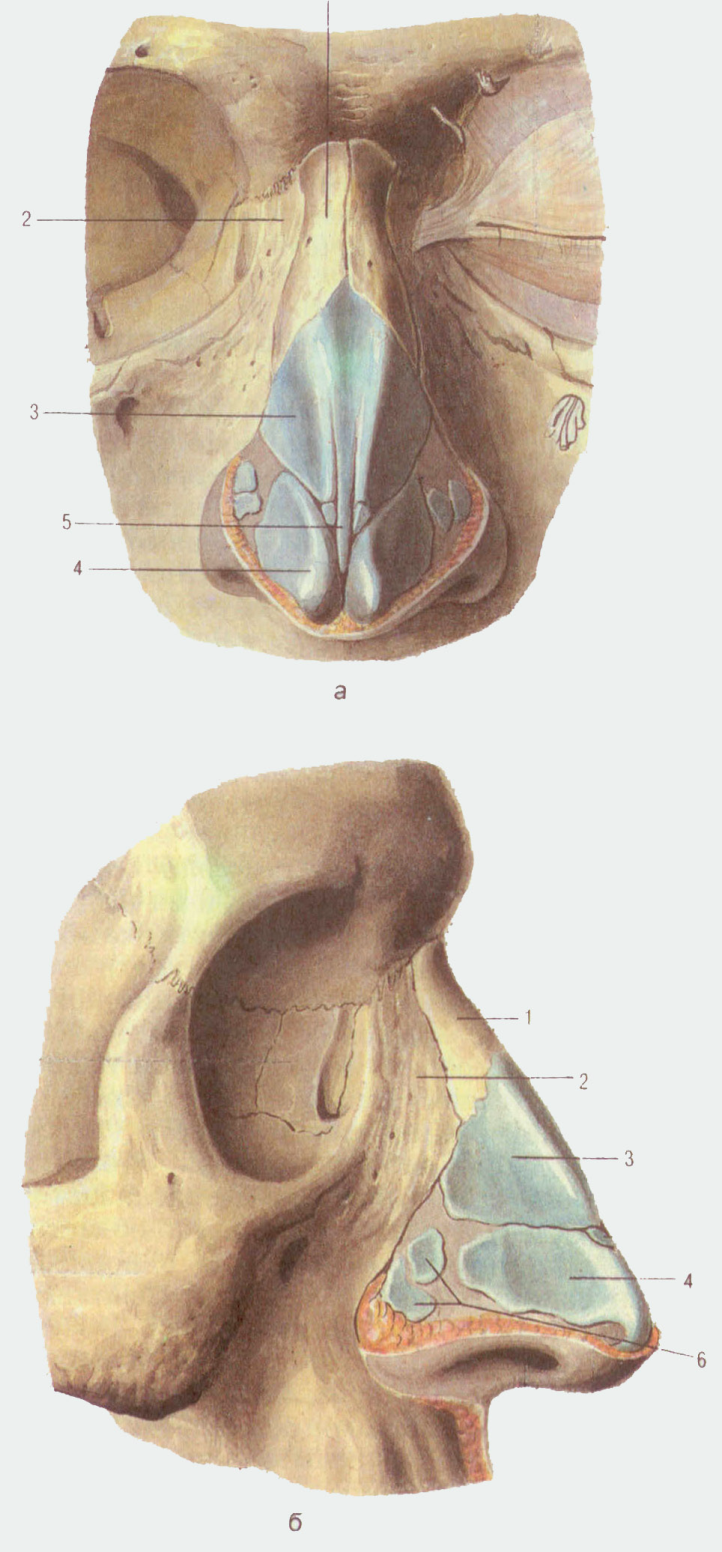 А при третьей изгиб перегородки практически полностью перекрывает носовой ход.
А при третьей изгиб перегородки практически полностью перекрывает носовой ход.
Что делают при искривлении носовой перегородки
Искривление носовой перегородки нельзя исправить консервативными способами. И промывания, ингаляции, закапывания лекарственных препаратов и другие методики являются лишь симптоматическими методиками лечения. Они помогают уменьшить выраженность неприятных симптомов и на время справиться с вторичными инфекционно-воспалительными заболеваниями носа, придаточных пазух, слуховой трубы и среднего уха.
Операция на перегородке носа – это способ радикального решения проблемы. Задачами хирургического вмешательства является восстановление дыхания по обоим носовым ходам и устранение блока устьев околоносовых пазух.
При резекции носовой перегородки удаляется ее искривленная часть и оставляется опора для спинки носа. При сложных и обширных сочетанных деформациях могут дополнительно проводить пластику хряща с нанесением на него насечек, репозицией краев резецированных тканей и наложением швов.
Резекция носовой перегородки: разновидности и техника
Классическим видом хирургического лечения является резекция носовой перегородки по Киллиану. Существуют и авторские вариации этой эндоназальной операции.
Доступ к хрящу при резекции носовой перегородки подслизистый. Для этого после выполнения дугообразного разреза слизистую оболочку отодвигают от места операции. Это позволяет сохранить ее функциональную активность и сокращает период восстановления. Обнаженный хрящ иссекают. При резекции носа по Киллиану образовавшийся дефект («окно») прикрывают лишь листком слизистой оболочки, поэтому такой способ называется окончатым. При обширном иссечении хряща спинка носа может потерять должную опору и в последующем подвергнуться деформации. Кроме того, классический вариант операции является достаточно травматичным.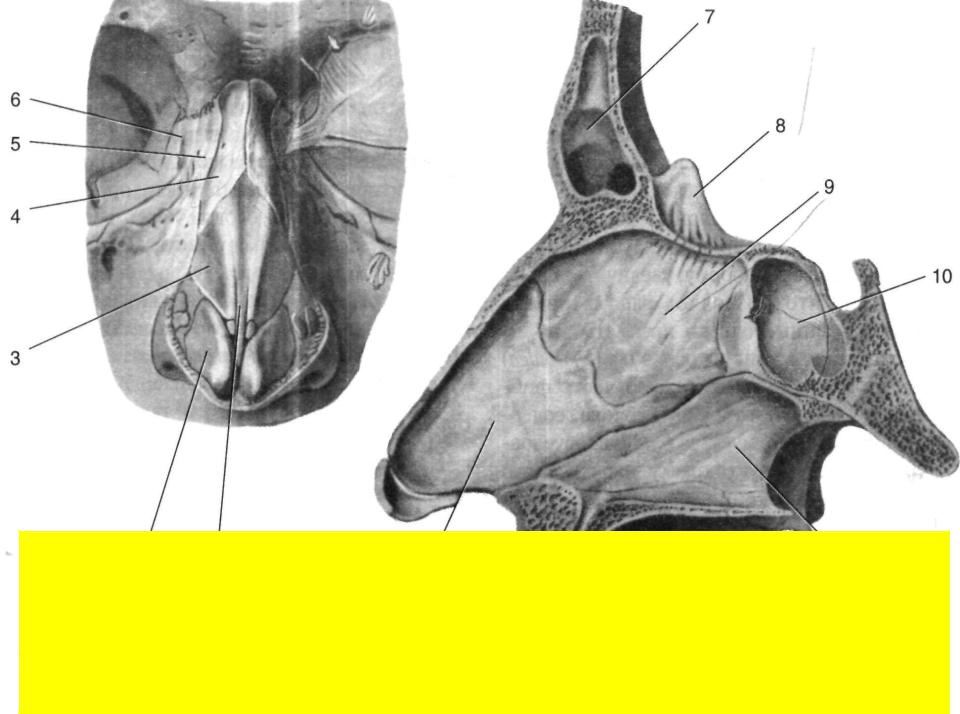
Используются и другие, более консервативные методики. К ним относят, например, циркулярную подслизистую резекцию носовой перегородки по Воячеку. При этом на хрящ наносятся несколько разрезов для обеспечения его подвижности в области искривления, после чего перегородку выпрямляют. Выбор техники операции зависит от навыков хирурга, характера и степени деформации. В настоящее время предпочтение отдается наименее травматичным методикам.
В клинике доктора Коренченко резекция носовой перегородки осуществляется эндоназально, с использованием современного высокотехнологичного оборудования. При необходимости в протокол операции включают вазотомию носовых раковин (редукцию носовых раковин). Для иссечения тканей опытные высококвалифицированные специалисты используют хирургический лазер и молекулярно-резонансный аппарат.
Это делает операцию малотравматичной и существенно снижает объем кровопотери. А восстановление тканей и устранение послеоперационного отека происходят максимально быстро. Такая резекция носовой перегородки не лишает пациента трудоспособности и проводится амбулаторно под местной анестезией.
Такая резекция носовой перегородки не лишает пациента трудоспособности и проводится амбулаторно под местной анестезией.
1.2. Этиология и патогенез / КонсультантПлюс
1.2. Этиология и патогенез
Основной причиной деформации перегородки носа (87%) являются ее травматические вывихи и переломы, в том числе в результате внутриутробной и родовой травмы, так как перегородка носа в этом возрасте состоит из нескольких незрелых хрящевых и несросшихся между собой легко травмирующихся отделов. По мере роста и формирования лицевого скелета в результате несоответствия роста костно-хрящевого остова перегородки носа и обрамляющей ее костной ее костной рамы, т.е. свода и дна носовой полости деформация перегородки носа становится более выраженной, вызывает стойкие функциональные нарушения и деформацию наружного носа.
Деформация перегородки носа также может быть обусловлена нарушением эмбриогенеза челюстно-лицевых зачатков, например, при врожденном незаращении твердого неба или аномалиях развития латеральной стенки полости носа.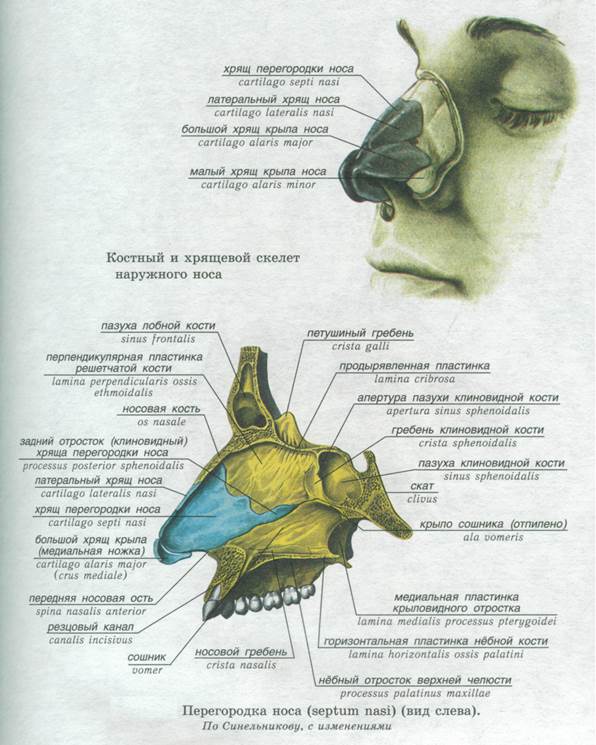 Значительно реже в детском возрасте наблюдаются вакатные деформации перегородки носа в результате полипоза и опухолей полости носа.
Значительно реже в детском возрасте наблюдаются вакатные деформации перегородки носа в результате полипоза и опухолей полости носа.
На основании анализа обстоятельств травмы можно выделить 5 типовых механизмов действия травмирующего агента, определяющих характерные виды переломов перегородки носа у детей.
При действии травмирующего агента спереди на кончик носа (9,3% случаев) чаще всего возникает вертикальный перелом четырехугольного хряща в каудальном отделе, сопровождающийся вывихом его из костного ложа в области носовой кости. Перегородка носа при этом деформируется в хрящевой части по типу шипа, сужая клапан носа. Костный скелет наружного носа при такой травме не повреждается, но с ростом лицевого скелета формируется деформация наружного носа в виде девиации его кончика в сторону.
В случае полного перелома хряща с ростом лицевого скелета хрящевой фрагмент в каудальном отделе отстает в развитии, а задние отделы хряща интенсивно растут в передне-верхнем направлении. Формируется так называемый двойной хрящ, который является частой интраоперационной находкой во время септопластики, выполняемой подросткам.
Формируется так называемый двойной хрящ, который является частой интраоперационной находкой во время септопластики, выполняемой подросткам.
Сильный удар по кончику носа нередко приводит к разрыву слизистой оболочки в области носогубной складки с выходом каудального отдела четырехугольного хряща в преддверие носа, при этом возможно образование вестибуло-назальных свищевых ходов, абсцессов перегородки и дна полости носа.
При действии травмирующего агента спереди на спинку носа (32,6% случаев) чаще всего происходит горизонтальный перелом четырехугольного хряща в передне-нижнем отделе с формированием деформации типа гребня. Исходная форма перегородки носа в значительной мере определяет виды деформации, которые наступают в результате травмы. Если перегородка носа ранее имела C-образный изгиб, то при ударе спереди или сверху она ломается в сторону изгиба. Если же она была в строго серединном положении, то происходит вывих хряща из костного ложа премаксиллы и сошника. Костная часть перегородки носа, как правило, не повреждается. При такой травме возможен перелом носовых костей без существенного смещения отломков и без деформации наружного носа.
При такой травме возможен перелом носовых костей без существенного смещения отломков и без деформации наружного носа.
При действии травмирующего агента сверху на спинку носа (6,2% случаев) в первую очередь образуются горизонтальные переломы в задних отделах костной части перегородки носа. Костные отломки перпендикулярной пластинки решетчатой кости и сошника заходят друг на друга, возможна травма ситовидной пластинки решетчатой кости, вызывающая носовую ликворею. Четырехугольный хрящ в зависимости от его исходной формы может надламываться, продолжая линию перелома костной части, либо изгибаться и вывихиваться из костного ложа на дне полости носа и соскальзывать с сошника. Образуется комбинированная костно-хрящевая деформация перегородки носа в виде гребня или деформация костной части перегородки носа в виде бокового смещения, существенно нарушающая респираторную функцию носа.
При такой травме всегда происходит перелом носовых костей, часто многооскольчатый, со смещением отломков кнутри. Возникает деформация наружного носа в виде западения и уплощения спинки. Если своевременно не выполнить репозицию отломков, то со временем формируется седловидная деформация носа, устраняемая ринопластикой.
Возникает деформация наружного носа в виде западения и уплощения спинки. Если своевременно не выполнить репозицию отломков, то со временем формируется седловидная деформация носа, устраняемая ринопластикой.
Действие травмирующего агента сбоку на спинку носа отмечается в 16,3% случаев, возникает в подавляющем большинстве у мальчиков после удара кулаком по носу во время драки. При этом отмечается перелом носовых костей и лобных отростков верхней челюсти с формированием деформацией наружного носа в виде западения одного ската (чаще левого) или бокового смещения всей спинки носа в сторону, устраняемых репозицией.
В ряде случаев происходит комбинированный перелом ячеек решетчатой кости и медиальной стенки орбиты. Рентгенологически при этом определяются кровоизлияния в полости решетчатой кости, реже — в орбиту. Клинически в таких случаях может определяться подкожная эмфизема в области медиального угла глаза и нижнего века.
При таком механизме травмы происходит горизонтальный перелом перегородки носа в верхних отделах, чаще без смещения, реже — с образованием угловой деформации костной части.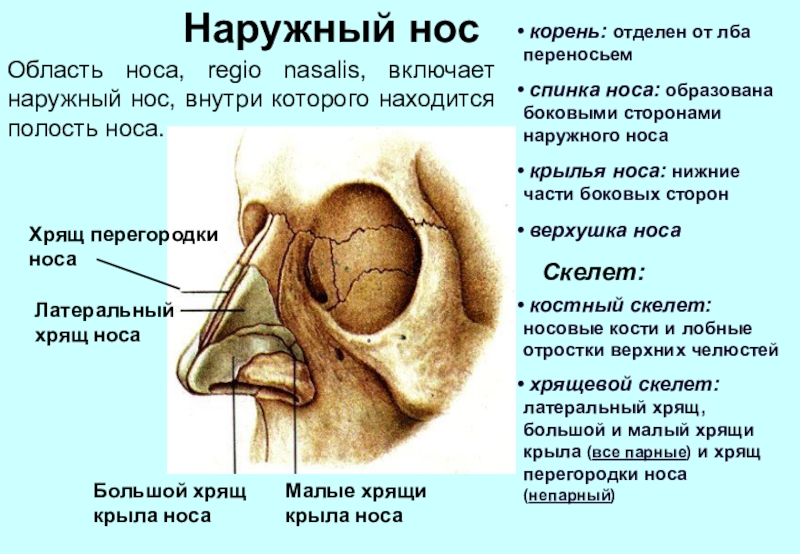 Поскольку эти деформации располагаются в верхних отделах полости носа, то серьезных нарушений респираторной функции носа они не вызывают, но в ряде случаев нарушается обоняние. Четырехугольный хрящ, как правило, не ломается, а лишь S- или C-образно изгибается.
Поскольку эти деформации располагаются в верхних отделах полости носа, то серьезных нарушений респираторной функции носа они не вызывают, но в ряде случаев нарушается обоняние. Четырехугольный хрящ, как правило, не ломается, а лишь S- или C-образно изгибается.
Травматические повреждения, возникающие при ударе лицом о твердую поверхность (6,2% случаев) возникают в результате автотравмы или падения с высоты, отличаются разнообразием клинических комбинаций и тяжестью общего состояния пострадавшего, т.к. сопровождаются сочетанными повреждениями других частей тела.
Повреждения перегородки характеризуются множественными разнонаправленными переломами, определяющими сложные комбинированные деформации костно-хрящевой части. Наиболее характерной при такой травме является сложная комбинированная деформация (так называемая «смятая» перегородка), обусловленная множественными вертикально направленными переломами.
При таком механизме травмы переломы костей и перегородки носа часто комбинируются с переломами стенок лобных и верхнечелюстных пазух, кровоизлияниями в них, с ранениями лица и повреждениями внутриносовых структур, переломами основания черепа и внутричерепными гематомами.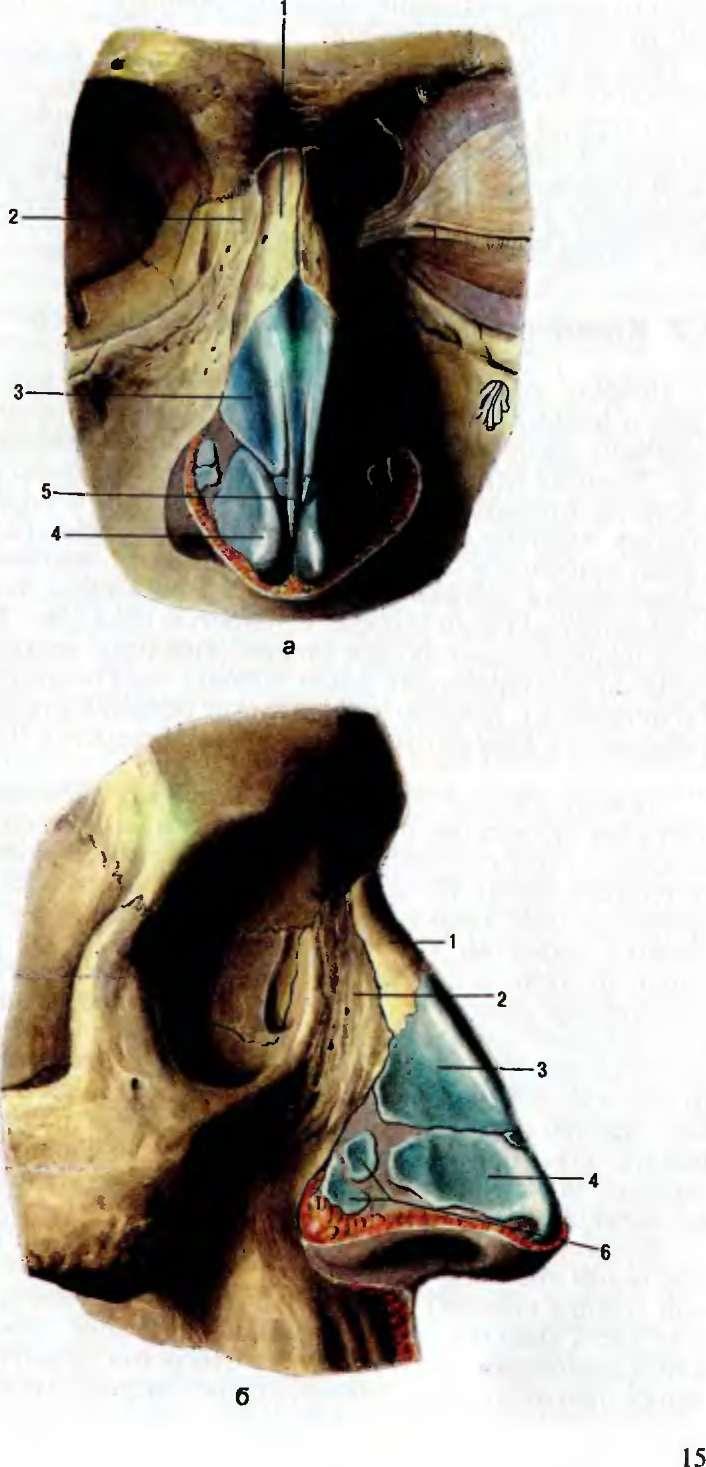
Таким образом, несмотря на многочисленные клинические варианты травматических деформаций перегородок носа, имеются определенные закономерности смещения отломков в зависимости от механизма травмы, уточнить которые удается в 75,2% случаев. Знание этих особенностей помогает планировать использование тех или иных оперативно-технических способов хирургического лечения [1, 2, 3, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15].
Операции на носовой перегородке — цены на лечение, консультация у ведущих специалистов
В зависимости от степени искривления методики проведения операций различаются.
При выраженных (как правило посттравматических) деформациях проводится септопластика (исправление перегородки) с ремоделированием: хрящ полностью выделяется из костного ложа, исправляется до правильной анатомической формы, после чего устанавливается на прежнее место. При необходимости устанавливаются поддерживающие сплиты – силиконовые вставки, фиксирующие хрящ в правильном положении до полного заживления. Длительность такой операции составляет как правило 1-1,5 часа, пребывание в стационаре — 2 суток.
Длительность такой операции составляет как правило 1-1,5 часа, пребывание в стационаре — 2 суток.
При незначительных искривлениях хрящ полностью не извлекается, эндоскопическим способом точечно исправляются только изменённые участки. Длительность операции 30 мин – 1 час., пребывание в стационаре – 2 суток.
При необходимости при проведении обеих операций может исп…
Реабилитационный период занимает как правило ококоло 3-4 недель. В это время очень важно строго соблюдать все рекомендации врача – любые отклонения от назначений могут привести к таким грозным осложнениям, как дегенерация (отмирание) слизистой полости носа, перфорации (образования отверстия) перегородки. Наиболее опасным осложнением является профузное (обильное) носовое кровотечение, остановить которое возможно только в условиях стационара.
Однако при соблюдении врачебных рекомендаций и благоприятном исходе все риски окупаются сторицей: качество жизни пациентов после операции существенно улучшается – исчезает постоянное ощущение заложенности носа, дыхание становится легким и свободным, практически мгновенно уходят хронические ЛОР-заболевания, восстанавливается нормальный сон, нормализуется давление.
Москва, улица Бакунинская, дом 1-3
Ближайшая станция метро: Бауманская
Белорусский государственный медицинский университет
1.ЖЕЛЧНЫЙ ПУЗЫРЬ
, vesica biliaris (fellea). Грушевидной формы образование, длиной 8 — 12 см. Рис. А. 2.Дно желчного пузыря
, fundus ves. biliaris. Округлой формы cлепой конец желчного пузыря, направленный каудально. Рис. А. 3.Тело желчного пузыря
, corpus ves.biliaris. Находится между дном и шейкой. Рис. А. 4.Шейка желчного пузыря
, collum ves.biliaris. Продолжается в пузырный проток. Рис. А. 5.Серозная оболочка
, tuniсa serosa. Перитонеальный покров желчного пузыря. Рис. Б. 6.Подсерозная основа
, tela subserosa. Соединительная ткань, расположенная под серозной оболочкой. Рис. Б. 7.Мышечная оболочка
, tunica muscularis. Образована неисчерченными мышечными волокнами. Рис. Б. 8.
Рис. Б. 8.Слизистая оболочка
, tunica mucosa. Состоит из однослойного высокого цилиндрического эпителия. Рис. Б. 9.Складки слизистой оболочки
, plicae mucosae. Выступают в просвет пузыря, в результате чего его поверхность приобретает ячеистый вид. Рис. А, Рис. Б. 10.Пузырный проток
, ductus cysticus. Выводной проток желчного пузыря, который соединяется с общим печеночным протоком и образует общий желчный проток. Рис. А. 11.Спиральная складка
, plica spiralis. Находится в шейке желчного пузыря и пузырном протоке. Рис. А. 12.Общий желчный проток
, ductus choledochus (biliaris). Образуется из общего печеночного и пузырного протоков. Открывается на большом сосочке двенадцатиперстной кишки. Рис. А. 13.Сфинктер общего желчного протока
, m.sphincter ductus choledochi. Утолщение кругового слоя мышечной оболочки. Расположен перед печеночноподжелудочной ампулой. Рис. А. 14.Печеночно-поджелудочная ампула
, ampulla hepatopancreatica.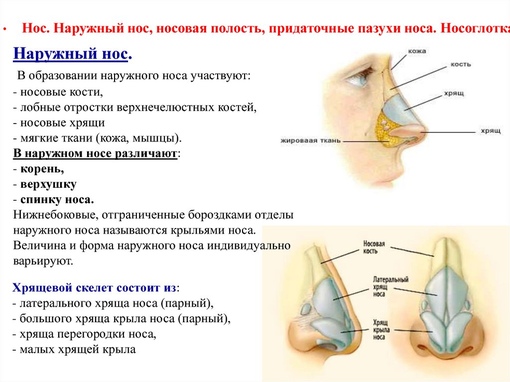 Расширение в стенке двенадцатиперстной кишки после соединения общего желчного протока и ductus pancreaticus. Рис. А. 15.
Расширение в стенке двенадцатиперстной кишки после соединения общего желчного протока и ductus pancreaticus. Рис. А. 15.Сфинктер печеночно-поджелудочной ампулы (сфинктер ампулы)
, m. sphincter ampullae hepatopancreaticae (m. sphincter ampullae). Утолщение циркулярного слоя мышечной оболочки вокруг ампулы (сфинктер Одди). Рис. А. 16.Железы желчных протоков
, gll. mucosae biliares. Имеют трубчато-альвеолярное строение и расположены в слизистой оболочке. Рис. А.17.
ДЫХАТЕЛЬНЫЙ АППАРАТ (ДЫХАТЕЛЬНАЯ СИСТЕМА)
, apparatus respiratorius (systema respiratorium). 18.НАРУЖНЫЙ НОС
, nasus externus. Рис. Д. 19.Корень носа
, radix nasi (nasalis). Расположен в верхней части лица, между глазницами. Рис. Г, Рис. Д. 20.Спинка носа
, dorsum nasi. Рис. Д. 21.Верхушка носа
, apex nasi. Рис. Д. 22.Крылья носа
, alae nasi. Нижними краями ограничивают ноздри. Рис. Д. 23.
Рис. Д. 23.Хрящи носа
, cartilaginеs nasi (nasalis). Формируют хрящевой скелет носа. Рис. В. 24. [Латеральный хрящ носа
, cartilago nasi lateralis]. Долгое время рассматривался как часть перегородки носа, а не как две самостоятельные хрящевые пластинки, каждая из которых частично срастается с cartilago septi nasi. Рис. В. 25.Большой хрящ крыла
, cartilago alaris major. Имеет форму крючка. Окружает ноздри и формирует верхушку носа. Рис. В. 26.Медиальная ножка
,crus mediale. Образует передненижнюю часть хрящевой перегородки носа. Рис. В, Рис. Г. 27.Латеральная ножка
, crus laterale. Окружает ноздрю с латеральной стороны. Рис. В. 28.Малые хрящи крыльев
, cartilagines alares minores. Обособленные хрящевые пластинки небольших размеров, дополняющие большой хрящ носа. Рис. В.29.
Добавочные носовые хрящи
, cartilagines nasales accessoriae. Расположены между хрящом перегородки носа и большим хрящом крыла. Встречаются непостоянно. 30.
Встречаются непостоянно. 30.Хрящ перегородки носа,
cartilago septi nasi. Самостоятельная широкая хрящевая пластинка в составе перегородки носа между перпендикулярной пластинкой решетчатой кости и сошником. Рис. Г. 30а.Латеральный отросток
, processus lateralis. Отходит от заднелатерального края хряща перегородки носа и срастается с латеральным хрящом носа. Рис. Г. 31.Задний (клиновидный) отросток
, processus posterior (sphenoidalis). Расположен между сошником и перпендикулярной пластинкой os sphenoidale. Иногда доходит до клиновидной кости. Рис. Г. 32.Сошниково-носовой [[Якобсона]] хрящ
, cartilago vomeronasalis. [[Jacobson]]. Узкая хрящевая полоска, которая находится между нижним фрагментом cartilago septi nasi и сошником. Рис. Г.33. [[
Подвижная часть перегородки носа
, pars mobilis septi nasi]]. Передненижняя часть перегородки носа, образованная медиальной ножкой большого хряща крыла.
Хрящ из носа заменил хрящ в колене
Клетки из носовой перегородки сформировали новый хрящ для коленного сустава и подавили остеоартрозное воспаление в больном колене.
Боль и опухание суставов могут быть признаком остеоартроза, когда разрушается суставной хрящ. Причин у остеоартроза много, от врождённых мутаций в белках хряща до травм, ожирения и изменений в гормональном фоне. Но в любом случае дегенерация хряща сопровождается воспалением. Это воспаление пытаются подавить, но если ничего не помогает, остаётся только ставить искусственный сустав. Но искусственные суставы изнашиваются, их нужно менять, что опять же не слишком удобно, особенно, если проблема с суставом случилась в относительно молодом возрасте.
Сотрудники Базельского университета предлагают иной способ избавиться от остеоартроза — заменить больной хрящ в суставе на хрящ из носовой перегородки. Конечно, носовая перегородка маленькая, а хрящ в коленном суставе — большой. Но в хрящах живут клетки соединительной ткани, которые размножаются и формируют хрящ — выделяют белки и другие молекулы, которые заполняют межклеточное пространство и образуют ткань.
Но в хрящах живут клетки соединительной ткани, которые размножаются и формируют хрящ — выделяют белки и другие молекулы, которые заполняют межклеточное пространство и образуют ткань.
Вот такие клетки, взятые из носовой перегородки, в специальной питательной среде сделали кусок нового хряща, который потом пересадили в коленный сустав. Эксперименты ставили с животными, сначала с мышами, а потом с овцами с остеоартритными суставами. Выращенный хрящ должен был не просто выдержать механическое воздействие — он должен быть устойчив к воспалению.
В статье в Science Translational Medicine говорится, что носовой хрящ, оказавшись в остеоартритном колене, успешно подавлял воспаление. Когда клетки носового хряща оказывались в коленном суставе, то здесь слабели воспалительные молекулярные сигнальные цепочки, которые обычно активны при остеоартрите. Авторы работы объясняют это тем, что клетки суставного хряща и клетки из носовой перегородки, хотя и занимаются, казалось бы, одним и тем же делом, во многом отличаются друг от друга. В частности, в ходе эмбрионального развития те и другие образуются от разных клеток-предшественников. И там, где клетки суставного хряща ничего не могут сделать с воспалением, клетки хряща носовой перегородки успешно его подавляют.
В частности, в ходе эмбрионального развития те и другие образуются от разных клеток-предшественников. И там, где клетки суставного хряща ничего не могут сделать с воспалением, клетки хряща носовой перегородки успешно его подавляют.
После успешных экспериментов на животных исследователи проделали ту же операцию с двумя больными с тяжёлым остеоартритом, которым уже ничего не оставалось, кроме как ставить искусственный сустав. У обоих утихли боли в колене, и само колено стало работать лучше. То, что сустав пошёл на поправку, было видно и по магнитно-резонансной томографии. В перспективе ещё предстоит выяснить, подходит ли такой способ лечения ко всем случаям остеоартрита. Но, так или иначе, учитывая распространённость болезни, для многих людей такая замена хряща могла бы стать настоящим спасением.
Что такое хрящ перегородки?
Хрящ — это тип ткани, которая мягче и гибче, чем кость, но тверже мышечной ткани. Хрящ перегородки делит нижнюю часть носа на две части или ноздри. Перегородочный хрящ человека является важным элементом как внешнего вида, так и нормального дыхания.
Перегородочный хрящ человека является важным элементом как внешнего вида, так и нормального дыхания.
Хрящ перегородки простирается от кончика носа до середины носа. Над этим хрящом находятся носовые кости, которые легче всего ощущаются на переносице, где обычно располагаются очки. Носовые кости соединяются с хрящом перегородки около середины носа. Около вершины носовых полостей перпендикулярная пластина этикоидной кости прикрепляется к хрящу. Вместе эти кости и хрящи помогают определить форму носа.
Два четырехугольных хряща лежат по обе стороны от хряща перегородки, около средней части носа. Они известны как верхние боковые хрящи, и они прикрепляются к носовым костям вблизи верха и сливаются с перегородочным хрящом вокруг середины носа. Если травма приводит к отделению этих хрящей от костей, дыхание может быть затруднено.
Форма нижней части носа в значительной степени определяется нижними боковыми хрящами. Они лежат под хрящом перегородки и слабо связаны на каждой стороне носа. Нижние боковые хрящи различаются по толщине, размеру и форме, создавая различные кончики носа, практически гарантируя, что у двух людей не будет одинаковых носов.
Нижние боковые хрящи различаются по толщине, размеру и форме, создавая различные кончики носа, практически гарантируя, что у двух людей не будет одинаковых носов.
Когда хрящ перегородки становится изогнутым или не исправляется после травмы, может возникнуть состояние, известное как отклоненная перегородка. Отклонение не может заметно повлиять на внешний вид, но оно может повлиять на воздушный поток через носовые проходы. Храп является одним из возможных последствий отклоненной перегородки. Хирургия, как правило, является предпочтительным методом лечения для достижения постоянного решения для искривленного или искривленного перегородочного хряща.
Хрящ перегородки также может быть поврежден опухолями, язвами или травмой, которая вызывает отверстие или перфорацию в перегородке. Люди, которые подвергаются воздействию сварочных паров в течение длительного периода времени, а также те, кто злоупотребляет определенными запрещенными наркотиками, также подвержены риску заболевания. Перфорированная перегородка обычно вызывает свистящий звук, когда пациент дышит, и может возникнуть боль или кровотечение. Если перфорация не заживает сама по себе, может потребоваться хирургическое вмешательство для восстановления поврежденной перегородки.
Перфорированная перегородка обычно вызывает свистящий звук, когда пациент дышит, и может возникнуть боль или кровотечение. Если перфорация не заживает сама по себе, может потребоваться хирургическое вмешательство для восстановления поврежденной перегородки.
ДРУГИЕ ЯЗЫКИ
границ | Искривление носовой перегородки как следствие контролируемых BMP изменений свойств хряща
Введение
Хрящ носовой перегородки, который делит носовую полость на две части, представляет собой гиалиновый хрящ. Он противостоит деформации, обеспечивает структурную поддержку средней части лица и является стимулятором роста средней зоны лица. Искривление носовой перегородки (NSD), которое описывает непрямую, деформированную носовую перегородку, является распространенной аномалией и может наблюдаться у 80% населения (Serifoglu et al., 2017). NSD может быть незаметным, но часто связан с различной степенью обструкции носовых дыхательных путей, приводящей к нарушению дыхания во сне (SDB) (Wang et al. , 2016; Alsufyani et al., 2017; Mandour et al., 2019). Врожденно-приобретенное отклонение часто развивается во время быстрого роста средней части лица и связано с замедлением роста средней части лица и сильной предрасположенностью к SDB (D’Ascanio et al., 2010). Другой распространенной этиологией является травма лица и носа. Для корректирующей септопластики часто требуется хрящевой трансплантат, который в зависимости от места забора либо ограничен в количестве, либо связан с заболеваемостью донорского участка, либо и то, и другое.Несмотря на возможность моделирования результатов хирургической септопластики практически до операции (Moghaddam et al., 2020), частота ревизий остается высокой и превышает 15% (Lavernia et al., 2019). Такой высокий показатель может быть следствием нашего ограниченного понимания этиологии педиатрической врожденной приобретенной НСД и связанных с ней вторичных осложнений (Baddam et al., 2020). В настоящее время неизвестно, имеет ли НСД основную молекулярную и клеточную этиологию, хотя это может быть связано с врожденными случаями, которые связаны с уменьшением роста средней части лица.
, 2016; Alsufyani et al., 2017; Mandour et al., 2019). Врожденно-приобретенное отклонение часто развивается во время быстрого роста средней части лица и связано с замедлением роста средней части лица и сильной предрасположенностью к SDB (D’Ascanio et al., 2010). Другой распространенной этиологией является травма лица и носа. Для корректирующей септопластики часто требуется хрящевой трансплантат, который в зависимости от места забора либо ограничен в количестве, либо связан с заболеваемостью донорского участка, либо и то, и другое.Несмотря на возможность моделирования результатов хирургической септопластики практически до операции (Moghaddam et al., 2020), частота ревизий остается высокой и превышает 15% (Lavernia et al., 2019). Такой высокий показатель может быть следствием нашего ограниченного понимания этиологии педиатрической врожденной приобретенной НСД и связанных с ней вторичных осложнений (Baddam et al., 2020). В настоящее время неизвестно, имеет ли НСД основную молекулярную и клеточную этиологию, хотя это может быть связано с врожденными случаями, которые связаны с уменьшением роста средней части лица.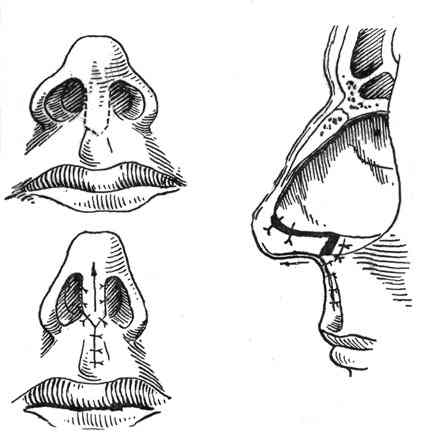
Хотя хрящ носовой перегородки является гиалиновым хрящом, он отличается от других гиалиновых хрящей по нескольким аспектам. Во-первых, это активный центр роста средней части лица, и большая часть его роста происходит, по крайней мере, у мышей, за счет гипертрофии хондроцитов (Baddam et al., 2021b). Во-вторых, гипертрофированные хондроциты, расположенные в средней части перегородки, не подвергаются апоптозу или окостенению, за исключением задней части, формирующей костную перпендикулярную пластинку решетчатой кости (Wealthall and Herring, 2006).В-третьих, это зеркальная структура с клетками-предшественниками и незрелыми хондроцитами, расположенными по обе стороны перегородки, близко к надхрящнице (Baddam et al., 2021b). В-четвертых, гипертрофированные хондроциты окружены плотным матриксом коллагеновых фибрилл, который, как считается, обеспечивает жесткость и жесткость. В какой степени любая из этих особенностей изменена при NSD, остается неизученным.
Изменения гиалинового хряща в контексте патологий хряща лучше всего изучены при остеоартрите (ОА) коленного сустава. Там общепринятыми признаками являются потеря гликозаминогликанов (GAG), увеличение гипертрофии хондроцитов наряду с увеличением количества коллагена X (COL X), Indian Hedgehog (IHH) и транскрипционного фактора 2, связанного с Runt (RUNX2), увеличение количества активных форм кислорода. (АФК) и увеличение метаболизма глюкозы (Четина, Маркова, 2018). Другие изменения, такие как измененная эластичность, остаются спорными, так как в одних исследованиях отмечается снижение эластичности, а в других предполагается увеличение (Coles et al., 2010; Кандела и др., 2016). На молекулярном уровне при ОА коленного сустава можно наблюдать нарушения в нескольких сигнальных сетях. Описано снижение количества антагонистов WNT Dickkopf1 (DKK1) и родственного белка Frizzled (FRZB) одновременно с увеличением нефосфорилированного бета-катенина (NPBC) в качестве показателя канонической передачи сигналов WNT (Liu et al., 2016; Zhong et al. ., 2016б). Изменения в передаче сигналов костного морфогенетического белка (BMP) связаны с гипертрофией хондроцитов.
Там общепринятыми признаками являются потеря гликозаминогликанов (GAG), увеличение гипертрофии хондроцитов наряду с увеличением количества коллагена X (COL X), Indian Hedgehog (IHH) и транскрипционного фактора 2, связанного с Runt (RUNX2), увеличение количества активных форм кислорода. (АФК) и увеличение метаболизма глюкозы (Четина, Маркова, 2018). Другие изменения, такие как измененная эластичность, остаются спорными, так как в одних исследованиях отмечается снижение эластичности, а в других предполагается увеличение (Coles et al., 2010; Кандела и др., 2016). На молекулярном уровне при ОА коленного сустава можно наблюдать нарушения в нескольких сигнальных сетях. Описано снижение количества антагонистов WNT Dickkopf1 (DKK1) и родственного белка Frizzled (FRZB) одновременно с увеличением нефосфорилированного бета-катенина (NPBC) в качестве показателя канонической передачи сигналов WNT (Liu et al., 2016; Zhong et al. ., 2016б). Изменения в передаче сигналов костного морфогенетического белка (BMP) связаны с гипертрофией хондроцитов. BMP7 подавляет, тогда как BMP2 способствует гипертрофии хондроцитов и деградации матрикса (Caron et al., 2013), а снижение уровня BMP7 напрямую связано с ОА (Merrihew et al., 2003; Huang et al., 2018). Неизвестно, происходят ли и в какой степени подобные изменения в хряще искривленной перегородки.
BMP7 подавляет, тогда как BMP2 способствует гипертрофии хондроцитов и деградации матрикса (Caron et al., 2013), а снижение уровня BMP7 напрямую связано с ОА (Merrihew et al., 2003; Huang et al., 2018). Неизвестно, происходят ли и в какой степени подобные изменения в хряще искривленной перегородки.
Мы недавно описали, что у мышей со специфичной для нервного гребня делецией BMP7 (BMP7 ncko ) развивается гипоплазия средней части лица и обструкция носовых дыхательных путей (Baddam et al., 2021a). Отличительной чертой этой модели является развитие значительного отклонения носовой перегородки в юношеском возрасте, что связано с прогрессированием аномального дыхания.Делеция BMP7 из развивающихся конечностей приводит к дегенерации суставного хряща и синовиальному воспалению (Abula et al., 2015). Содержание протеогликанов и экспрессия аггрекана были снижены, тогда как экспрессия матриксной металлопротеиназы-13 (ММР13) увеличилась. В этом исследовании мы задались вопросом, связаны ли подобные молекулярные и клеточные изменения, наблюдаемые в коленном суставе, с развитием NSD у мышей BMP7 ncko .
Мы демонстрируем, что BMP7 экспрессируется в надхрящнице и носовых хондроцитах на протяжении всего постнатального развития.Потеря BMP7 приводит к гистоморфологическим изменениям к 4-недельному возрасту, когда устанавливается NSD. Сопоставив эти изменения с молекулярными изменениями, используя анализ экспрессии генов, количественную протеомику дробовика и анализ иммунофлуоресценции в различные моменты времени развития, мы определили, что изменения свойств хондроцитов предшествуют NSD. Это включало приобретение маркеров эластичного хряща, переключение на метаболизм глюкозы, а также увеличение молекулярных маркеров, обычно связанных с ОА коленного сустава.Потеря BMP7 также была связана со значительным увеличением канонической передачи сигналов WNT в зрелых хондроцитах. Сопутствующее снижение BMP2 у мышей BMP7 ncko восстановило изменение передачи сигналов WNT, предотвратило развитие отклонения и избавило от гипоплазии средней части лица, демонстрируя, что баланс BMP2 и BMP7 синергически определяет свойства хряща. Поскольку многие клеточные и молекулярные изменения при NSD у мышей BMP7 ncko имеют патофизиологическое сходство с ОА коленного сустава, это исследование создает прецедент для необходимости дальнейшего понимания свойств носового хряща для использования в тканевой инженерии или клинических применениях, связанных с регенерацией. повреждения хряща при ОА коленного сустава.
Поскольку многие клеточные и молекулярные изменения при NSD у мышей BMP7 ncko имеют патофизиологическое сходство с ОА коленного сустава, это исследование создает прецедент для необходимости дальнейшего понимания свойств носового хряща для использования в тканевой инженерии или клинических применениях, связанных с регенерацией. повреждения хряща при ОА коленного сустава.
Материалы и методы
Модели животных
В этом исследовании использовались как самцы, так и самки мышей. Все мыши содержались на фоне C57BL/6 и подвергались обратному скрещиванию в течение не менее 10 поколений. Экспрессия BMP7 была идентифицирована с использованием репортерных мышей BMP7LacZ (Malik et al., 2020). Мышей BMP7 fl/fl , также называемых мышами BMP7 ctrl , скрещивали с мышами WNT1-cre для удаления BMP7 из клеток нервного гребня (впоследствии называемых мышами BMP7 ncko ) (мыши Malik 9014et) др. , 2020). Для спасения NSD, мышей BMP7 NCKO NCKO были скрещены с BMP2 WT / FL (Malik et al., 2018) Для получения BMP2 WT / FL BMP7 NCKO , также упоминается до BMP2 het BMP7 ncko впоследствии. Отслеживание линии проводилось с использованием мышей mT/mG [ Gt(ROSA)26Sor TM 4(ACTB-tdTomato,-EGFP)Luo /J] (Muzumdar et al., 2007). Экспрессию BMP7 и идентификацию клеток нервного гребня в носовой перегородке проводили при рождении (0-й день после рождения, P0), через 2 недели (14-й день после рождения, P14) и через 4 недели (30-й день после рождения, P30).Кроме того, носовую перегородку мышей BMP7 ctrl и BMP7 ncko также оценивали в указанные выше моменты времени. Мышей BMP2 het BMP7 ncko оценивали только на P30.
, 2020). Для спасения NSD, мышей BMP7 NCKO NCKO были скрещены с BMP2 WT / FL (Malik et al., 2018) Для получения BMP2 WT / FL BMP7 NCKO , также упоминается до BMP2 het BMP7 ncko впоследствии. Отслеживание линии проводилось с использованием мышей mT/mG [ Gt(ROSA)26Sor TM 4(ACTB-tdTomato,-EGFP)Luo /J] (Muzumdar et al., 2007). Экспрессию BMP7 и идентификацию клеток нервного гребня в носовой перегородке проводили при рождении (0-й день после рождения, P0), через 2 недели (14-й день после рождения, P14) и через 4 недели (30-й день после рождения, P30).Кроме того, носовую перегородку мышей BMP7 ctrl и BMP7 ncko также оценивали в указанные выше моменты времени. Мышей BMP2 het BMP7 ncko оценивали только на P30.
Микрокомпьютерная томография (мкКТ) Анализ
Морфологические изменения носовой перегородки BMP7 ctrl и BMP7 ncko ( n = 3/генотип) оценивали на Р14 и Р30 с помощью микроКТ. Получение и реконструкция носовой перегородки с использованием MILabs μCT в Школе стоматологии Университета Альберты проводились, как описано ранее (Baddam et al., 2021b). Степень и тяжесть NSD были количественно определены на коронарных репрезентациях носовой перегородки с использованием программного обеспечения Amira. Два ориентира были размещены на абатменте, где перпендикулярная пластина встречается с решетчатой пластиной решетчатой кости и где сошник сочленяется с небной костью, соответственно. Между этими двумя точками была проведена прямая линия, обозначающая гипотетическую прямую перегородку, а ее длина принималась за ее высоту (а).Затем фактическая длина перегородки была определена путем отслеживания фактической длины перегородки (b). Степень отклонения определяли как (б – а)/а и выражали в процентах. Продольные эксперименты для определения того, становится ли NSD тяжелым с течением времени, оценивали путем расчета степени NSD на одних и тех же мышах дважды в двух разных возрастах [4 недели (P30) и 10 недель (P74)].
Получение и реконструкция носовой перегородки с использованием MILabs μCT в Школе стоматологии Университета Альберты проводились, как описано ранее (Baddam et al., 2021b). Степень и тяжесть NSD были количественно определены на коронарных репрезентациях носовой перегородки с использованием программного обеспечения Amira. Два ориентира были размещены на абатменте, где перпендикулярная пластина встречается с решетчатой пластиной решетчатой кости и где сошник сочленяется с небной костью, соответственно. Между этими двумя точками была проведена прямая линия, обозначающая гипотетическую прямую перегородку, а ее длина принималась за ее высоту (а).Затем фактическая длина перегородки была определена путем отслеживания фактической длины перегородки (b). Степень отклонения определяли как (б – а)/а и выражали в процентах. Продольные эксперименты для определения того, становится ли NSD тяжелым с течением времени, оценивали путем расчета степени NSD на одних и тех же мышах дважды в двух разных возрастах [4 недели (P30) и 10 недель (P74)].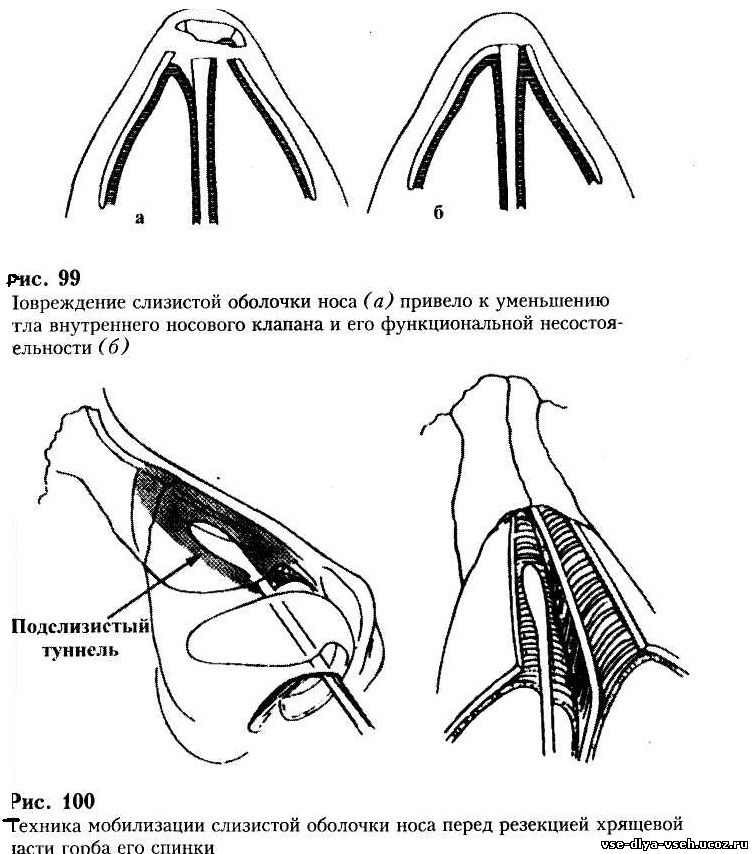
Секвенирование РНК
Тотальную РНК из изолированной носовой перегородки мышей P0 BMP7 ctrl и BMP7 ncko ( n = 4/генотип) экстрагировали реагентом TRIzol (Invitrogen).Библиотеки RNAseq были сконструированы из 500 нг тотальной РНК с использованием набора для подготовки библиотеки направленной РНК NEBNext Ultra II для компании Illumina (NEB). Полиаденилированные мРНК были обогащены олиго-dT, конъюгированными с парамагнитными шариками. Обогащенные мРНК фрагментировали химически и использовали для синтеза кДНК. кДНК подвергали репарации концов и А-хвосту, лигировали с линкерами и, наконец, индексировали с помощью ПЦР, чтобы обеспечить мультиплексирование во время секвенирования. Секвенирование проводили на приборе MiSeq в соответствии с протоколом парных концов 75 циклов.Необработанные данные секвенирования общедоступны на портале NCBI SRA под регистрационным номером PRJNA622501. Был проведен биоинформатический анализ, при котором фрагменты были сопоставлены с базой данных кДНК человека (GRCh48) с использованием Kallisto (Bray et al. , 2016) со 100 перестановками во время псевдовыравнивания. Дифференциальный анализ экспрессии данных RNAseq был проведен с использованием негативных биномиальных обобщенных линейных моделей с пакетом edgeRR (Love et al., 2014). Различия в распространенности генов с исправленным значением p <0.05 и логарифмическое изменение ≤–2 или ≤2 считались дифференциально выраженными. Графики среднего дифференциала были созданы с помощью edgeR, а тепловые карты были созданы с использованием тепловой карты пакета R. Онтологии генов были идентифицированы с использованием консорциума генных онтологий PANTHER (Mi et al., 2019). Для представления были выбраны только значимые онтологии (FDR <0,05).
, 2016) со 100 перестановками во время псевдовыравнивания. Дифференциальный анализ экспрессии данных RNAseq был проведен с использованием негативных биномиальных обобщенных линейных моделей с пакетом edgeRR (Love et al., 2014). Различия в распространенности генов с исправленным значением p <0.05 и логарифмическое изменение ≤–2 или ≤2 считались дифференциально выраженными. Графики среднего дифференциала были созданы с помощью edgeR, а тепловые карты были созданы с использованием тепловой карты пакета R. Онтологии генов были идентифицированы с использованием консорциума генных онтологий PANTHER (Mi et al., 2019). Для представления были выбраны только значимые онтологии (FDR <0,05).
Протеомика дробовика
Носовая перегородка мышей P14 и P30 BMP7 ctrl и BMP7 ncko ( n = 4/генотип) выделяли и замораживали до обработки для протеомных экспериментов.Белковые лизаты получали путем лизиса ткани в буфере, содержащем 1% SDS, в 200 мМ HEPES (pH 8,0) и полном коктейле ингибиторов протеазы TM (MilliporeSigma, Оквилл, Онтарио, Канада). Образцы восстанавливали 10 мМ дитиотреитола (ДТТ) в течение 30 мин при 37°С. После охлаждения при комнатной температуре (КТ) алкилирование цистеина осуществляли путем инкубации с конечной концентрацией 15 мМ иодацетамида в течение 25 мин в темноте при КТ. Затем образцы осаждали смесью ацетон/метанол, трижды промывали метанолом и трипсинизировали (Trypsin Gold от Promega, Мэдисон, Висконсин, США).pH доводили до 6,0 с помощью HCl. Для мечения пептидных α- и ε-аминов образцы инкубировали в течение 18 ч при 37°С с изотопно-тяжелыми [40 мМ 13CD2O + 20 мМ NaBh4CN (цианоборогидрид натрия)] или легкими метками [40 мМ легкий формальдегид (Ch3O) + 20 мМ NaBh4CN]. Затем образцы объединяли и подвергали хроматографии на С18, а затем подвергали жидкостной хроматографии и тандемной масс-спектрометрии.
Образцы восстанавливали 10 мМ дитиотреитола (ДТТ) в течение 30 мин при 37°С. После охлаждения при комнатной температуре (КТ) алкилирование цистеина осуществляли путем инкубации с конечной концентрацией 15 мМ иодацетамида в течение 25 мин в темноте при КТ. Затем образцы осаждали смесью ацетон/метанол, трижды промывали метанолом и трипсинизировали (Trypsin Gold от Promega, Мэдисон, Висконсин, США).pH доводили до 6,0 с помощью HCl. Для мечения пептидных α- и ε-аминов образцы инкубировали в течение 18 ч при 37°С с изотопно-тяжелыми [40 мМ 13CD2O + 20 мМ NaBh4CN (цианоборогидрид натрия)] или легкими метками [40 мМ легкий формальдегид (Ch3O) + 20 мМ NaBh4CN]. Затем образцы объединяли и подвергали хроматографии на С18, а затем подвергали жидкостной хроматографии и тандемной масс-спектрометрии.
Высокоэффективная жидкостная хроматография (ВЭЖХ) и масс-спектрометрия (МС)
Эксперименты по жидкостной хроматографии и масс-спектрометрии были проведены на основной установке масс-спектрометрии Южной Альберты (SAMS) в Университете Калгари, Канада.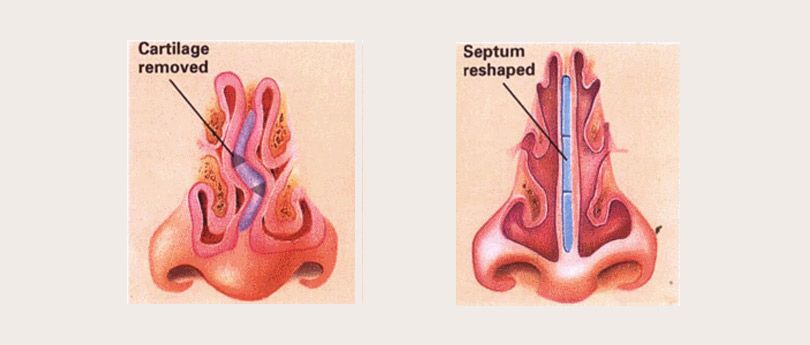 Анализ проводили на масс-спектрометре Orbitrap Fusion Lumos Tribrid (Thermo Scientific), работающем с Xcalibur (версия 4.0.21.10) и соединенном с системой Thermo Scientific Easy-nLC (нанопоточная жидкостная хроматография) 1200. Триптические пептиды (2 мкг) наносили на колонку C18 (75 мкм × 2 см; Acclaim PepMap 100, P/N 164946; Thermo Scientific) при скорости потока 2 мкл/мин растворителя А (0,1% муравьиной кислоты и 3 % ацетонитрила в воде чистоты ЖХ-МС). Затем пептиды подвергали электрораспылению с использованием напряжения 2,3 кВ в трубке для переноса ионов (300°C) устройства Orbitrap Lumos, работающего в положительном режиме.Сначала Orbitrap выполнил полное МС сканирование с разрешением 120 000 fwhm, чтобы обнаружить ион-предшественник, имеющий m/z от 375 до 1575 и заряд от +2 до +7. АРУ Orbitrap (автоматическая регулировка усиления) и максимальное время ввода были установлены на 4 × 10 5 и 50 мс соответственно. Orbitrap работал в режиме максимальной скорости с временем цикла 3 с для выбора прекурсора.
Анализ проводили на масс-спектрометре Orbitrap Fusion Lumos Tribrid (Thermo Scientific), работающем с Xcalibur (версия 4.0.21.10) и соединенном с системой Thermo Scientific Easy-nLC (нанопоточная жидкостная хроматография) 1200. Триптические пептиды (2 мкг) наносили на колонку C18 (75 мкм × 2 см; Acclaim PepMap 100, P/N 164946; Thermo Scientific) при скорости потока 2 мкл/мин растворителя А (0,1% муравьиной кислоты и 3 % ацетонитрила в воде чистоты ЖХ-МС). Затем пептиды подвергали электрораспылению с использованием напряжения 2,3 кВ в трубке для переноса ионов (300°C) устройства Orbitrap Lumos, работающего в положительном режиме.Сначала Orbitrap выполнил полное МС сканирование с разрешением 120 000 fwhm, чтобы обнаружить ион-предшественник, имеющий m/z от 375 до 1575 и заряд от +2 до +7. АРУ Orbitrap (автоматическая регулировка усиления) и максимальное время ввода были установлены на 4 × 10 5 и 50 мс соответственно. Orbitrap работал в режиме максимальной скорости с временем цикла 3 с для выбора прекурсора. Наиболее интенсивные ионы-предшественники, представляющие пептидный изотопный профиль и имеющие порог интенсивности не менее 5000, были выделены с использованием квадруполя и фрагментированы с помощью HCD (30% энергии столкновения) в мультиполе маршрутизации ионов.Ионы-фрагменты (MS2) анализировали в ионной ловушке при высокой скорости сканирования. АРУ и максимальное время инжекции были установлены на 1 × 10 4 и 35 мс соответственно для ионной ловушки. Динамическое исключение было включено на 45 с, чтобы избежать получения одного и того же иона-предшественника, имеющего сходное значение m/z (±10 м.д.).
Наиболее интенсивные ионы-предшественники, представляющие пептидный изотопный профиль и имеющие порог интенсивности не менее 5000, были выделены с использованием квадруполя и фрагментированы с помощью HCD (30% энергии столкновения) в мультиполе маршрутизации ионов.Ионы-фрагменты (MS2) анализировали в ионной ловушке при высокой скорости сканирования. АРУ и максимальное время инжекции были установлены на 1 × 10 4 и 35 мс соответственно для ионной ловушки. Динамическое исключение было включено на 45 с, чтобы избежать получения одного и того же иона-предшественника, имеющего сходное значение m/z (±10 м.д.).
Протеомные данные и анализ биоинформатики
Спектральные данные были сопоставлены с последовательностями пептидов в базе данных мышиных белков UniProt с использованием алгоритма Andromeda (Cox et al., 2011), реализованный в пакете программного обеспечения MaxQuant (Cox and Mann, 2008) v.1.6.0.1, при частоте ложных обнаружений совпадения пептидного спектра (FDR) <0,01. Параметры поиска включали допуск по массе 20 ppm для родительского иона, 0,5 Да для иона-фрагмента, карбамидометилирование остатков цистеина (+57,021464 Да), переменную модификацию N-конца путем ацетилирования (+42,010565 Да) и переменную степень окисления метионина (+ 15,994915 Да). N-концевое и тяжелое (+34,063116 Да) и легкое (+28,031300 Да) диметилирование лизина определяли как метки для относительного количественного определения.Специфичность сайта расщепления была установлена на трипсин/P, при этом допускалось до двух пропущенных расщеплений. Значимые пороговые значения выбросов были определены после преобразования log(2) с помощью анализа «ящик и ус» с использованием инструмента BoxPlotR (Altman and Krzywinski, 2016). Для количественного определения использовали минимум два различных пептида на белок. После анализа межквартильных диаграмм для анализа рассматривались белки с коэффициентом количественного определения ниже 0,65 и выше 1,5. Протеомные данные были переданы в Консорциум ProteomeXchange через партнерский репозиторий PRIDE с идентификатором набора данных px-submission PXD024813.
Параметры поиска включали допуск по массе 20 ppm для родительского иона, 0,5 Да для иона-фрагмента, карбамидометилирование остатков цистеина (+57,021464 Да), переменную модификацию N-конца путем ацетилирования (+42,010565 Да) и переменную степень окисления метионина (+ 15,994915 Да). N-концевое и тяжелое (+34,063116 Да) и легкое (+28,031300 Да) диметилирование лизина определяли как метки для относительного количественного определения.Специфичность сайта расщепления была установлена на трипсин/P, при этом допускалось до двух пропущенных расщеплений. Значимые пороговые значения выбросов были определены после преобразования log(2) с помощью анализа «ящик и ус» с использованием инструмента BoxPlotR (Altman and Krzywinski, 2016). Для количественного определения использовали минимум два различных пептида на белок. После анализа межквартильных диаграмм для анализа рассматривались белки с коэффициентом количественного определения ниже 0,65 и выше 1,5. Протеомные данные были переданы в Консорциум ProteomeXchange через партнерский репозиторий PRIDE с идентификатором набора данных px-submission PXD024813. Список всех дифференциально регулируемых белков представлен в дополнительных таблицах 1, 2.
Список всех дифференциально регулируемых белков представлен в дополнительных таблицах 1, 2.
Экстракция РНК Количественная полимеразная цепная реакция в реальном времени (qRT-PCR)
Носовую перегородку мышей P0, P14 и P30 BMP7 ctrl и BMP7 ncko ( n = 3/генотип) вырезали и обрабатывали для выделения РНК. Экстракцию РНК и qRT-PCR проводили, как описано ранее (Malik et al., 2020). Различие в кратности определяли в отношении гена домашнего хозяйства 36B4 с использованием метода ΔΔCt (Malik et al., 2020). Анализ проводился на биологической и технической трехкратной повторности. Показанные данные являются представителями трех биологических образцов каждого генотипа. Список пар праймеров, используемых для анализа экспрессии генов, приведен в дополнительной таблице 3.
Обработка тканей и гистология
Мышей BMP7 ctrl и BMP7 ncko ( n = 3/генотип/возраст) фиксировали с использованием 4% параформальдегида (PFA). Черепа мышей вскрывали и декальцинировали в 0.5М этилендиаминтетрауксусная кислота (ЭДТА), как описано ранее (Malik et al., 2020). Черепа мышей фронтально заливали парафином и делали срезы с использованием микротома 820 Spencer при 7 микронах. Для гистологического окрашивания срезы депарафинизировали, как описано ранее (Baddam et al., 2021b).
Черепа мышей вскрывали и декальцинировали в 0.5М этилендиаминтетрауксусная кислота (ЭДТА), как описано ранее (Malik et al., 2020). Черепа мышей фронтально заливали парафином и делали срезы с использованием микротома 820 Spencer при 7 микронах. Для гистологического окрашивания срезы депарафинизировали, как описано ранее (Baddam et al., 2021b).
Гистологические красители
Медиальные срезы носовой перегородки использовались для гистологического и иммунофлуоресцентного анализа (Baddam et al., 2021b). Окрашивание гематоксилином и эозином (H&E) и сафранином O на носовой перегородке мышей P14 и P30 BMP7 ctrl и BMP7 ncko мышей ( n = 3/генотип/возраст) проводили, как описано ранее. др., 2020; Баддам и др., 2021b). Окрашивание пикросириусом проводили с использованием протокола окрашивания Sirius Red от IHC world (Kiernan, 2001). H&E выполняли для оценки общей морфологии. Окрашивание сафранином О использовали для окрашивания хряща, а окрашивание пикросириусом красным использовали для окрашивания коллагеновых волокон. Изображения были получены на микроскопе Olympus IX73 с использованием объективов 20X и 40X.
Изображения были получены на микроскопе Olympus IX73 с использованием объективов 20X и 40X.
Иммунофлуоресценция
Иммунофлуоресцентное окрашивание было проведено на медиальных носовых перегородках Парафиновые участки P0, P14, P30 BMP7 CTRL и BMP7 NCKO и P30 BMP2 BMP7 NCKO MICE как описанные ранее (Baddam и другие., 2021б). Спецификации первичных и вторичных антител представлены в дополнительной таблице 4. Изображения были получены на микроскопе Olympus IX73 с использованием 20-кратных объективов. Анализ проводили в трех биологических повторностях. Ранее были продемонстрированы отрицательные контроли, демонстрирующие фоновое окрашивание (Baddam et al., 2021b).
Окрашивание LacZ
репортерных мыши P0, P14 и P30 BMP7LacZ ( n = 3/генотип/возраст) окрашивали с использованием ранее описанной процедуры окрашивания (Malik et al., 2020). Черепа мышей фиксировали, декальцинировали, обрабатывали, как описано выше.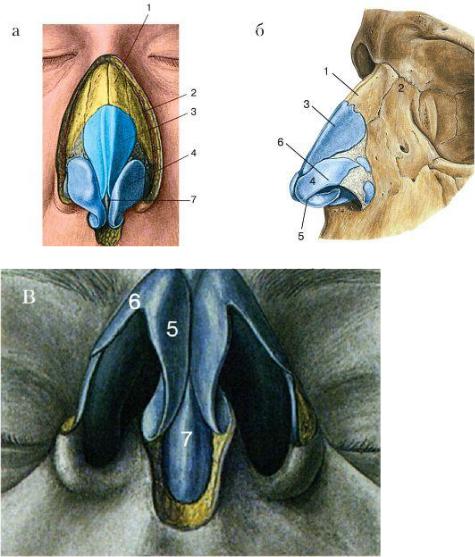 Парафиновые срезы контрастировали с использованием протокола окрашивания сафранином О.
Парафиновые срезы контрастировали с использованием протокола окрашивания сафранином О.
Статистика
На графиках показаны отдельные измерения, а также среднее значение ± стандартное отклонение, если применимо. Двусторонний независимый тест t был проведен для проверки статистической значимости между мышами BMP7 ctrl и BMP7 ncko . Microsoft Excel использовался для отображения графиков, и был проведен независимый непарный тест t с использованием статистического онлайн-программного обеспечения (Socscistatistics, 2020). ∗ P < 0,05; ∗∗ P < 0,01; ∗∗∗ P < 0,001; нс, несущественно P > 0,05.
Утверждение исследования
Это исследование было одобрено Комитетом медицинских наук по уходу за животными и их использованию Университета Альберты (протокол №: AUP1149). Все эксперименты в этом исследовании проводились в соответствии с рекомендациями Канадского совета по уходу за животными.
Результаты
BMP7 экспрессируется в носовом хряще, и его делеция приводит к искривлению носовой перегородки
Передний и медиальный срезы носовой перегородки использовались для гистологической оценки носовой перегородки (Baddam et al., 2021б). Изображения были получены в месте примыкания носовой перегородки к внутреннему шву, а также в середине носовой перегородки (рис. 1А). Чтобы определить, когда и где BMP7 экспрессируется в носовой перегородке, мы использовали репортерных мышей BMP7LacZ ( n = 3/возраст). Экспрессия наблюдалась на абатменте во всех исследованных возрастных группах [рис. 1A’–C; Послеродовой день 0 (P0), Послеродовой день 14 (P14), Послеродовой день 30 (P30)]. В средней области перегородки BMP7 четко экспрессировался в хондроцитах, а также в перихондральной выстилке, начиная с P0 (рис. 1D-F).Экспрессия менялась с течением времени, при этом наименьшая экспрессия наблюдалась в хондроцитах на P14 (рис. 1E). Отслеживание клонов подтвердило происхождение нервного гребня хондроцитов перегородки и надхрящницы ( n = 3/возраст) (рис. 1G-I). Как было показано ранее (Baddam et al., 2021a), носовая перегородка у мышей BMP7 ncko сильно искривляется на P30 (рис. 1J–O) ( n = 3/возраст/генотип) со степенью отклонения достигая от 10 до 35% (рис. 1L; p < 0.001) и варьирует по степени тяжести с течением времени (рис. 10). Носовая перегородка у мышей быстро растет между P7-P14 и P21-P30, но показывает значительное уменьшение между P14 и P21 (Vora et al., 2015; Baddam et al., 2021b). Поскольку у мышей BMP7 ncko в дополнение к NSD развивается гипоплазия средней части лица, мы предположили, что изменения внешнего вида и свойств хряща связаны с развитием этих двух патологий.
1G-I). Как было показано ранее (Baddam et al., 2021a), носовая перегородка у мышей BMP7 ncko сильно искривляется на P30 (рис. 1J–O) ( n = 3/возраст/генотип) со степенью отклонения достигая от 10 до 35% (рис. 1L; p < 0.001) и варьирует по степени тяжести с течением времени (рис. 10). Носовая перегородка у мышей быстро растет между P7-P14 и P21-P30, но показывает значительное уменьшение между P14 и P21 (Vora et al., 2015; Baddam et al., 2021b). Поскольку у мышей BMP7 ncko в дополнение к NSD развивается гипоплазия средней части лица, мы предположили, что изменения внешнего вида и свойств хряща связаны с развитием этих двух патологий.
Рисунок 1. Делеция BMP7 в клетках нервного гребня приводит к искривлению носовой перегородки (NSD). (A) мкКТ-реконструкция черепа мыши P30, демонстрирующая расположение носовой перегородки, откуда были получены изображения. Окрашенные LacZ (синий) парафиновые срезы носовой перегородки, демонстрирующие экспрессию BMP7 у репортерных мышей P0 (A’, D) , P14 (B, E) и P30 (C, F) Bmp7LacZ.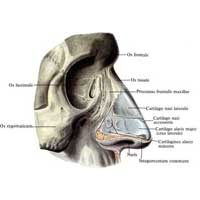 Клетки нервного гребня в носовой перегородке P0 (G) , P14 (H) и P30 (I) , идентифицированные антителом к зеленому флуоресцентному белку (GFP) (красное). DAPI используется для окрашивания ядер (серый).Масштабная линейка = 50 мкм. Трехмерный вид спереди мышей P30 Bmp7 ctrl (J) и Bmp7 ncko (K) , демонстрирующий искривление носовой перегородки, обведенное белым цветом. (L) Количественная оценка NSD у мышей P14 и P30 ( n = 4/генотип/возраст). Двухмерное фронтальное поперечное сечение носовой перегородки у мышей Bmp7 ctrl (M) и Bmp7 ncko (N) . (O) Количественная оценка отклонения носовой перегородки у мышей Bmp7 ncko с течением времени ( n = 3/возраст).Точки данных представляют отдельных мышей с планками погрешностей, демонстрирующими биологическую изменчивость. ins, внутренний шов; ns, носовая перегородка; р, надхрящница; я, резец; м, нижняя челюсть.
Клетки нервного гребня в носовой перегородке P0 (G) , P14 (H) и P30 (I) , идентифицированные антителом к зеленому флуоресцентному белку (GFP) (красное). DAPI используется для окрашивания ядер (серый).Масштабная линейка = 50 мкм. Трехмерный вид спереди мышей P30 Bmp7 ctrl (J) и Bmp7 ncko (K) , демонстрирующий искривление носовой перегородки, обведенное белым цветом. (L) Количественная оценка NSD у мышей P14 и P30 ( n = 4/генотип/возраст). Двухмерное фронтальное поперечное сечение носовой перегородки у мышей Bmp7 ctrl (M) и Bmp7 ncko (N) . (O) Количественная оценка отклонения носовой перегородки у мышей Bmp7 ncko с течением времени ( n = 3/возраст).Точки данных представляют отдельных мышей с планками погрешностей, демонстрирующими биологическую изменчивость. ins, внутренний шов; ns, носовая перегородка; р, надхрящница; я, резец; м, нижняя челюсть. P14, постнатальный день 14; P30, постнатальный день 30; P74, постнатальный день 74. *** p < 0,001, двусторонний независимый t -критерий. н.с., не имеет значения. Масштабная линейка (A–F) : 20 мкм и (G, H) : 50 мкм.
P14, постнатальный день 14; P30, постнатальный день 30; P74, постнатальный день 74. *** p < 0,001, двусторонний независимый t -критерий. н.с., не имеет значения. Масштабная линейка (A–F) : 20 мкм и (G, H) : 50 мкм.
Апоптоз предшествует искривлению носовой перегородки, и в искривленной перегородке наблюдается снижение содержания ГАГ
На P14 гистологический анализ носовой перегородки с использованием H&E, Safranin O и Picrosirius Red показал сравнимую клеточную организацию с округлыми (зрелыми/гипертрофическими) клетками, ограниченными средними и плоскими (незрелыми/предшественниками хондроцитов) клетками, расположенными на периферии (рис. 2А; n = 3/генотип).На P30 хрящ BMP7 ncko потерял эту четкую организацию. Степень гипертрофии уменьшилась, на периферии стало меньше плоских клеток. Окрашивание сафранином О уменьшилось, что свидетельствует об уменьшении ГАГ. Отложение коллагеновых волокон было асимметричным с повышенным отложением на внешней стороне отклонения и уменьшением на противоположной стороне (рис. 2В, n = 3/генотип). Клеточная пролиферация (PCNA) не пострадала. Апоптоз, измеренный окрашиванием расщепленной каспазы 3, увеличивался только на P14 без видимых различий на P30.Таким образом, вспышка клеточного апоптоза предшествует клеточной дезорганизации и уменьшению ГАГ, что указывает на то, что изменения свойств хондроцитов могут предрасполагать к развитию НСД.
2В, n = 3/генотип). Клеточная пролиферация (PCNA) не пострадала. Апоптоз, измеренный окрашиванием расщепленной каспазы 3, увеличивался только на P14 без видимых различий на P30.Таким образом, вспышка клеточного апоптоза предшествует клеточной дезорганизации и уменьшению ГАГ, что указывает на то, что изменения свойств хондроцитов могут предрасполагать к развитию НСД.
Рисунок 2. Гибель клеток изменила организацию гликозаминогликанов и коллагеновых фибрилл у мышей Bmp7 ncko . Гистологическое окрашивание парафиновых срезов носовой перегородки в моменты времени P14 (A) и P30 (B) ( n = 3/генотип/возраст).Окрашивание гематоксилином (фиолетовый) и эозином (розовый) (H&E) для общей морфологии носовой перегородки. Репрезентативное окрашивание сафранином O (красный) для хрящей и окрашивание пикросириусом красным (розовато-красный) для коллагеновых волокон. Снижение окрашивания сафранином О наблюдалось только у мышей P30 Bmp7 ncko . Для выявления пролиферации в носовой перегородке было проведено иммунофлуоресцентное окрашивание с использованием антитела к PCNA (красный цвет) со срезами, контрастно окрашенными DAPI для ядер (серый цвет). Никаких изменений в пролиферации не наблюдалось.Апоптоз был охарактеризован с использованием антител CASP3 (красный) и контрастно окрашен DAPI для ядер (серый). Апоптоз предшествует искривлению носовой перегородки у мышей Bmp7 ncko . PCNA, ядерный антиген пролиферирующих клеток; CASP3, расщепленная каспаза 3. Стрелки обозначают положительное окрашивание/сигнал. Масштабная линейка (гистологические пятна) = 20 мкм и (Immunos) = 50 мкм.
Для выявления пролиферации в носовой перегородке было проведено иммунофлуоресцентное окрашивание с использованием антитела к PCNA (красный цвет) со срезами, контрастно окрашенными DAPI для ядер (серый цвет). Никаких изменений в пролиферации не наблюдалось.Апоптоз был охарактеризован с использованием антител CASP3 (красный) и контрастно окрашен DAPI для ядер (серый). Апоптоз предшествует искривлению носовой перегородки у мышей Bmp7 ncko . PCNA, ядерный антиген пролиферирующих клеток; CASP3, расщепленная каспаза 3. Стрелки обозначают положительное окрашивание/сигнал. Масштабная линейка (гистологические пятна) = 20 мкм и (Immunos) = 50 мкм.
Белки, участвующие в организации внеклеточного матрикса и клеточном метаболизме, дерегулируются перед отклонением носовой перегородки
Мы выполнили протеомику «шотган» (рис. 3А) на изолированных P14 и P30 BMP7 ctrl и BMP7 ncko носовой перегородки ( n = 4/генотип/возраст изменений в носовой перегородке до мутантома). и после отклонения перегородки.База данных STRING использовалась для определения онтологии и картирования белок-белковых взаимодействий. На P14 было идентифицировано несколько кластеров, отражающих снижение количества белков, участвующих в организации внеклеточной структуры, клеточной адгезии и деградации внеклеточного матрикса (рис. 3B), на примере эластина (ELN), коллагена I (COL 1), коллагена II (COL 2). ), остеопонтин (SPP1) и аггрекан (ACAN). Некоторые из этих белков, а также сниженный mTOR связаны с сигнальным путем PI3K-AKT, что указывает на снижение этого важного пути, регулирующего пролиферацию, выживание и метаболизм клеток (Yu and Cui, 2016).В то же время активировался кластер белков, участвующих в передаче сигналов ретиноевой кислоты и метаболизме липидов, таких как CYP1A2 и ALDh2A2. Вместе это предполагает, что потеря BMP7 связана с изменением клеточного метаболизма на P14.
и после отклонения перегородки.База данных STRING использовалась для определения онтологии и картирования белок-белковых взаимодействий. На P14 было идентифицировано несколько кластеров, отражающих снижение количества белков, участвующих в организации внеклеточной структуры, клеточной адгезии и деградации внеклеточного матрикса (рис. 3B), на примере эластина (ELN), коллагена I (COL 1), коллагена II (COL 2). ), остеопонтин (SPP1) и аггрекан (ACAN). Некоторые из этих белков, а также сниженный mTOR связаны с сигнальным путем PI3K-AKT, что указывает на снижение этого важного пути, регулирующего пролиферацию, выживание и метаболизм клеток (Yu and Cui, 2016).В то же время активировался кластер белков, участвующих в передаче сигналов ретиноевой кислоты и метаболизме липидов, таких как CYP1A2 и ALDh2A2. Вместе это предполагает, что потеря BMP7 связана с изменением клеточного метаболизма на P14.
Рис. 3. Измененная организация внеклеточного матрикса (ECM) и клеточный метаболизм предшествуют искривлению носовой перегородки у мышей Bmp7 ncko . (A) Рабочий процесс протеомного эксперимента и анализа ( n = 4/возраст/генотип).Программное обеспечение STRINGdb использовалось для идентификации белок-белковых взаимодействий. (B) Дифференциально экспрессируемые белки и связанные с ними биологические процессы изменены у мышей P14 Bmp7 ncko по сравнению с Bmp7 ctrl . (C) Дифференциально экспрессируемые белки и связанные с ними биологические процессы изменены у мышей P30 Bmp7 ncko по сравнению с Bmp7 ctrl . * Обозначает белки с повышенной экспрессией у мышей Bmp7 ncko .Белки, участвующие в передаче сигналов ретиноевой кислоты и метаболизме липидов, активировались через 2 недели, тогда как белки, участвующие в метаболизме глюкозы, активировались через 4 недели. Список всех дифференциально регулируемых белков представлен в дополнительных таблицах 1, 2.
(A) Рабочий процесс протеомного эксперимента и анализа ( n = 4/возраст/генотип).Программное обеспечение STRINGdb использовалось для идентификации белок-белковых взаимодействий. (B) Дифференциально экспрессируемые белки и связанные с ними биологические процессы изменены у мышей P14 Bmp7 ncko по сравнению с Bmp7 ctrl . (C) Дифференциально экспрессируемые белки и связанные с ними биологические процессы изменены у мышей P30 Bmp7 ncko по сравнению с Bmp7 ctrl . * Обозначает белки с повышенной экспрессией у мышей Bmp7 ncko .Белки, участвующие в передаче сигналов ретиноевой кислоты и метаболизме липидов, активировались через 2 недели, тогда как белки, участвующие в метаболизме глюкозы, активировались через 4 недели. Список всех дифференциально регулируемых белков представлен в дополнительных таблицах 1, 2.
На П30, при установлении НСД, наблюдалась несколько иная картина изменений протеома (рис. 3В). У мышей BMP7 ncko наблюдали увеличение белков, участвующих в реакции на кислородсодержащие соединения, каталитическую активность и организацию цитоскелета, таких как декорин (DCN).Кроме того, наблюдались изменения процессинга мРНК и процессов метаболизма нуклеотидов. Интересно, что увеличение гексокиназы 1 (HK1), альдолазы A (ALDOA) и глицеральдегид-3-фосфатдегидрогеназы (GAPDH) указывало на увеличение гликолиза, и белки, участвующие в метаболизме РНК, такие как гетерогенный ядерный рибонуклеопротеин U (HNRPU), также были активны. регулируемый. Таким образом, мы определили глобальные динамические изменения в нескольких клеточных процессах в носовом хряще с мутацией BMP7, наиболее заметно внеклеточном матриксе и клеточном метаболизме как до, так и после установления NSD.Однако пространственная информация об экспрессии этих белков в перегородке неизвестна.
3В). У мышей BMP7 ncko наблюдали увеличение белков, участвующих в реакции на кислородсодержащие соединения, каталитическую активность и организацию цитоскелета, таких как декорин (DCN).Кроме того, наблюдались изменения процессинга мРНК и процессов метаболизма нуклеотидов. Интересно, что увеличение гексокиназы 1 (HK1), альдолазы A (ALDOA) и глицеральдегид-3-фосфатдегидрогеназы (GAPDH) указывало на увеличение гликолиза, и белки, участвующие в метаболизме РНК, такие как гетерогенный ядерный рибонуклеопротеин U (HNRPU), также были активны. регулируемый. Таким образом, мы определили глобальные динамические изменения в нескольких клеточных процессах в носовом хряще с мутацией BMP7, наиболее заметно внеклеточном матриксе и клеточном метаболизме как до, так и после установления NSD.Однако пространственная информация об экспрессии этих белков в перегородке неизвестна.
Увеличение гипертрофии хондроцитов и приобретение маркеров эластичного хряща в искривленной носовой перегородке BMP7
ncko Мыши Анализ экспрессии генов и иммунофлуоресцентное окрашивание ( n = 3/возраст/генотип) проводили с использованием антител против белков, идентифицированных в данных протеомики. Мы оценили экспрессию компонентов внеклеточного матрикса, связанных с гиалиновым хрящом, гипертрофией хряща и ОА на P0, P14, P30, чтобы зафиксировать время любых изменений.Мы дополнительно сравнили эти белковые изменения с изменениями в экспрессии генов (рис. 4А). COL II был значительно повышен на P0 BMP7 ncko мышей ( p <0,05), но стал сопоставим на P30. Экспрессия COL VI, гена перицеллюлярного матрикса (Zelenski et al., 2015), изначально была сопоставимой, но показала значительное снижение на P30 ( p <0,05). Однако экспрессия белка была увеличена в месте отклонения. OPN, ген, участвующий в раннем хондрогенезе (Gerstenfeld and Shapiro, 1996; Baddam et al., 2021b), был значительно снижен в носовой перегородке с мутацией P14. Кроме того, на P30 экспрессия белка OPN была потеряна в подмножестве клеток, которые, по-видимому, отходят от надхрящницы в месте отклонения. Таким образом, потеря BMP7 изменяет экспрессию нескольких компонентов внеклеточного матрикса, связанных с хондрогенезом.
Мы оценили экспрессию компонентов внеклеточного матрикса, связанных с гиалиновым хрящом, гипертрофией хряща и ОА на P0, P14, P30, чтобы зафиксировать время любых изменений.Мы дополнительно сравнили эти белковые изменения с изменениями в экспрессии генов (рис. 4А). COL II был значительно повышен на P0 BMP7 ncko мышей ( p <0,05), но стал сопоставим на P30. Экспрессия COL VI, гена перицеллюлярного матрикса (Zelenski et al., 2015), изначально была сопоставимой, но показала значительное снижение на P30 ( p <0,05). Однако экспрессия белка была увеличена в месте отклонения. OPN, ген, участвующий в раннем хондрогенезе (Gerstenfeld and Shapiro, 1996; Baddam et al., 2021b), был значительно снижен в носовой перегородке с мутацией P14. Кроме того, на P30 экспрессия белка OPN была потеряна в подмножестве клеток, которые, по-видимому, отходят от надхрящницы в месте отклонения. Таким образом, потеря BMP7 изменяет экспрессию нескольких компонентов внеклеточного матрикса, связанных с хондрогенезом.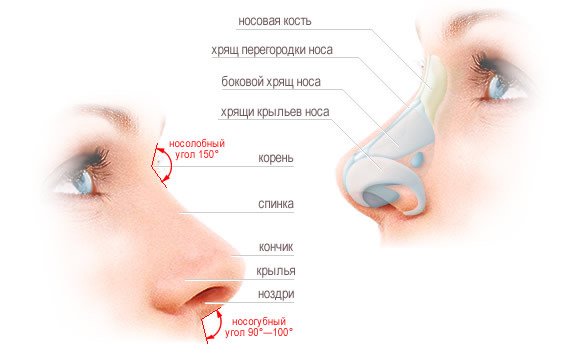
Рис. 4. Bmp7 ncko мыши подвергаются гипертрофии хондроцитов и приобретают маркеры эластичного хряща. (A) qRT-PCR рассеченной носовой перегородки, демонстрирующий дифференциальную экспрессию генов коллагена II (COL II), коллагена VI (COL VI) и остеопонтина (OPN) в моменты времени P0, P14 и P30.Иммунофлуоресцентное окрашивание COL II выявило увеличение на P0 (верхняя панель) и снижение на P30 (нижняя панель). Экспрессия COL VI была увеличена у мышей Bmp7 ncko , в то время как сниженная экспрессия OPN наблюдалась у мышей с мутацией P30 посредством иммунофлуоресценции. (B) Дифференциальная экспрессия генов, участвующих в гипертрофии хондроцитов RUNX2, коллагена X (COL X), индийского ежа (IHH), хрящевого кислого белка 1 (CRTAC1), остеокальцина и Bapx, что указывает на Bmp7 ncko мышей гипертрофия хондроцитов.Иммунофлюоресцентное окрашивание носовой перегородки P0 демонстрирует увеличение экспрессии RUNX2 в дополнение к увеличению экспрессии COL X и IHH у мышей P30 Bmp7 ncko . (C) Экспрессия эластина (ELN) наблюдалась в абатменте носовой перегородки и внутреннем шве (ins), а также в середине носовой перегородки у мышей P30 Bmp7 ncko . Экспрессия EMILIN3 также была значительно увеличена на P30 у мутантных мышей. Экспрессия декорина (DCN) и SERPINh2 (HSP47) также была идентифицирована как повышенная в мутантной носовой перегородке.Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: p < 0,05, **: p < 0,01, ***: p < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител. Масштабная линейка = 50 мкм.
(C) Экспрессия эластина (ELN) наблюдалась в абатменте носовой перегородки и внутреннем шве (ins), а также в середине носовой перегородки у мышей P30 Bmp7 ncko . Экспрессия EMILIN3 также была значительно увеличена на P30 у мутантных мышей. Экспрессия декорина (DCN) и SERPINh2 (HSP47) также была идентифицирована как повышенная в мутантной носовой перегородке.Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: p < 0,05, **: p < 0,01, ***: p < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител. Масштабная линейка = 50 мкм.
RUNX2 (Caron et al., 2013), COL X, IHH, CRTAC1 (Sanchez et al., 2017), Osteocalcin и BAPX (Caron et al., 2015) были связаны с гипертрофической дифференцировкой хондроцитов, при этом RUNX2, COL X и IHH являются наиболее часто оцениваемыми белками для гипертрофии хондроцитов.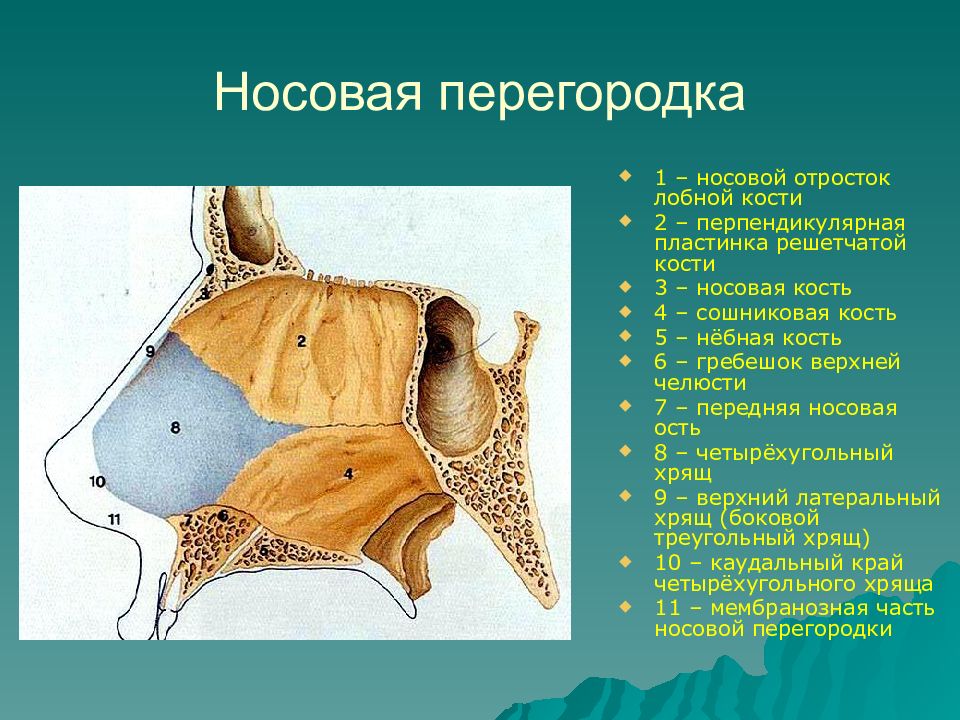 Хотя экспрессия гена RUNX2 (рис. 4B) была немного увеличена на P0 ( p <0,01), она не показала повышения, наблюдаемого у контрольных мышей на P14 ( p <0,01). Мыши BMP7 ncko указывают на раннюю индукцию гипертрофии хондроцитов. Окрашивание антителами к RUNX2 на P0 подтвердило повышенную экспрессию.В то время как экспрессия COL X была сопоставима на всех стадиях, экспрессия IHH была значительно увеличена ( p <0,05) в мутантной перегородке на P30. Окрашивание антителами показало увеличение COL X и IHH в искривленной перегородке. Экспрессия как CRTAC1, гена, участвующего в дифференцировке хондроцитов (Sanchez et al., 2017), так и BAPX, ингибитора гипертрофии хондроцитов (Caron et al., 2015), значительно повышалась на P14 и P30 ( ∗ p). < 0,05, ∗∗∗ р < 0.001). Остеокальцин, ранее описанный как накопление в начале гипертрофии (Lian et al., 1993), был значительно снижен на P14 ( p <0,001), но увеличился на P30 ( p <0,05) у мышей BMP7 ncko .
Хотя экспрессия гена RUNX2 (рис. 4B) была немного увеличена на P0 ( p <0,01), она не показала повышения, наблюдаемого у контрольных мышей на P14 ( p <0,01). Мыши BMP7 ncko указывают на раннюю индукцию гипертрофии хондроцитов. Окрашивание антителами к RUNX2 на P0 подтвердило повышенную экспрессию.В то время как экспрессия COL X была сопоставима на всех стадиях, экспрессия IHH была значительно увеличена ( p <0,05) в мутантной перегородке на P30. Окрашивание антителами показало увеличение COL X и IHH в искривленной перегородке. Экспрессия как CRTAC1, гена, участвующего в дифференцировке хондроцитов (Sanchez et al., 2017), так и BAPX, ингибитора гипертрофии хондроцитов (Caron et al., 2015), значительно повышалась на P14 и P30 ( ∗ p). < 0,05, ∗∗∗ р < 0.001). Остеокальцин, ранее описанный как накопление в начале гипертрофии (Lian et al., 1993), был значительно снижен на P14 ( p <0,001), но увеличился на P30 ( p <0,05) у мышей BMP7 ncko . Эти наблюдения показывают, что развитие NSD, по-видимому, связано с измененной регуляцией гипертрофии хондроцитов.
Эти наблюдения показывают, что развитие NSD, по-видимому, связано с измененной регуляцией гипертрофии хондроцитов.
Ключевой функцией гиалинового хряща является придание жесткости носовой полости. Учитывая значительные молекулярные изменения, которые предшествуют развитию отклонения, мы задались вопросом, может ли структурная деформация быть связана с приобретением маркеров эластичного хряща, которые обычно не наблюдаются в гиалиновом хряще.Иммунофлуоресцентное окрашивание на эластин (ELN), белок, содержащийся в большом количестве в эластичном хряще, показало, что потеря BMP7 приводила к экспрессии эластина в абатменте и средней области перегородки, что позволяет предположить, что по крайней мере части носовой перегородки действительно могли приобрести эластичный хрящ. свойства (рис. 4C). Это потенциальное переключение на маркеры эластичного хряща у мышей P30 BMP7 ncko было подтверждено увеличением экспрессии Emilin3 ( p <0,01), еще одного характерного гена эластичного хряща (Schiavinato et al. , 2012). Дополнительные доказательства структурных изменений и изменений коллагеновых фибрилл (рис. 2B) были получены в результате изменения распределения декорина (DCN) и HSP47 (Ishikawa et al., 2018) у мышей P30 BMP7 ncko .
, 2012). Дополнительные доказательства структурных изменений и изменений коллагеновых фибрилл (рис. 2B) были получены в результате изменения распределения декорина (DCN) и HSP47 (Ishikawa et al., 2018) у мышей P30 BMP7 ncko .
В совокупности наши данные свидетельствуют о том, что NSD связан с измененными свойствами хряща, в частности с усилением экспрессии маркеров эластичного хряща.
Увеличение гликолитической активности как следствие измененной передачи сигналов WNT и BMP
Анализ протеома на P30 выявил увеличение нескольких ключевых ферментов в пути гликолиза [(HK1, ALDOA, GAPDH и лактатдегидрогеназа B (LDHB)] (Lee et al., 2018) в мутантной перегородке (рис. 3С). Иммунофлуоресцентный анализ HK1 подтвердил повышенную экспрессию в искривленной носовой перегородке (рис. 5А). Белки, участвующие в метаболизме РНК, такие как HNRPU, также были идентифицированы (рис. 3C), и их повышенная экспрессия была подтверждена у мышей P30 BMP7 ncko . Путь гликолиза находится под контролем сигнальных путей WNT и BMP (Lee et al., 2018). Исследуя изменения в этих двух путях, мы выявили измененную экспрессию различных лигандов WNT (WNT3A, WNT6, WNT7A), антагонистов WNT (FRZB, DKK1), что приводит к усилению передачи сигналов WNT (NPBC) в зрелых хондроцитах мутантной перегородки P30 (рис. 5B). ).В целом изменения были наибольшими на P14. WNT3A, участвующий в канонической передаче сигналов WNT, был снижен на P14 ( p <0,001). Два других канонических лиганда WNT, WNT6 и WNT7A, активировались на P14 ( p <0,05) и P30 ( p <0,01) (Huang et al., 2018). WNT-антагонисты FRZB и DKK1 были активизированы и подавлены, соответственно, у мышей с мутацией P14 ( p <0,001) и в меньшей степени у P30 ( p <0,05). В целом, каноническая передача сигналов WNT, проявляющаяся окрашиванием ядер NPBC, была увеличена в мутантной перегородке, особенно в месте отклонения, тогда как окрашивание цитоплазмы NPBC наблюдалось у мышей BMP7 ctrl .
Путь гликолиза находится под контролем сигнальных путей WNT и BMP (Lee et al., 2018). Исследуя изменения в этих двух путях, мы выявили измененную экспрессию различных лигандов WNT (WNT3A, WNT6, WNT7A), антагонистов WNT (FRZB, DKK1), что приводит к усилению передачи сигналов WNT (NPBC) в зрелых хондроцитах мутантной перегородки P30 (рис. 5B). ).В целом изменения были наибольшими на P14. WNT3A, участвующий в канонической передаче сигналов WNT, был снижен на P14 ( p <0,001). Два других канонических лиганда WNT, WNT6 и WNT7A, активировались на P14 ( p <0,05) и P30 ( p <0,01) (Huang et al., 2018). WNT-антагонисты FRZB и DKK1 были активизированы и подавлены, соответственно, у мышей с мутацией P14 ( p <0,001) и в меньшей степени у P30 ( p <0,05). В целом, каноническая передача сигналов WNT, проявляющаяся окрашиванием ядер NPBC, была увеличена в мутантной перегородке, особенно в месте отклонения, тогда как окрашивание цитоплазмы NPBC наблюдалось у мышей BMP7 ctrl . Потеря BMP7 была связана со снижением MSX1, гена ниже по течению от передачи сигналов BMP, который способствует хондрогенной дифференцировке NCC (Méndez-Maldonado et al., 2018), как на P14 ( p <0,05), так и на P30 ( p ). <0,001) моменты времени (рис. 5C). Gremlin 1 (GREM1), антагонист BMP и ингибитор гипертрофии хондроцитов (Zhong et al., 2016a), значительно снижался после установления отклонения перегородки. Рецепторы BMP ALK2 и ALK6, ранее описанные для регуляции хондрогенной дифференцировки (Caron et al., 2013), были снижены на P14 (ALK2; p <0,001) и P30 (ALK6; p <0,01) соответственно.
Потеря BMP7 была связана со снижением MSX1, гена ниже по течению от передачи сигналов BMP, который способствует хондрогенной дифференцировке NCC (Méndez-Maldonado et al., 2018), как на P14 ( p <0,05), так и на P30 ( p ). <0,001) моменты времени (рис. 5C). Gremlin 1 (GREM1), антагонист BMP и ингибитор гипертрофии хондроцитов (Zhong et al., 2016a), значительно снижался после установления отклонения перегородки. Рецепторы BMP ALK2 и ALK6, ранее описанные для регуляции хондрогенной дифференцировки (Caron et al., 2013), были снижены на P14 (ALK2; p <0,001) и P30 (ALK6; p <0,01) соответственно.
Рисунок 5. Измененный метаболизм и передача сигналов Wnt у мышей P30 Bmp7 ncko . (A) Иммунофлуоресцентное окрашивание гексокиназы (HK1) и гетерогенного ядерного рибонуклеопротеина U (HNRPU) у мышей P30 Bmp7 ctrl и Bmp7 ncko выявило увеличение экспрессии (B) qRT-PCR рассеченной носовой перегородки в моменты времени P0, P14 и P30, демонстрирующие дифференциальную экспрессию генов лигандов Wnt (WNT3A, WNT6, WNT7A) и антагонистов Wnt (FRZB, DKK1). Иммуноокрашивание антагонистов Wnt (FRZB, DKK1) у мышей P30 Bmp7 ctrl и Bmp7 ncko демонстрирует увеличение экспрессии FRZB и снижение экспрессии DKK1 у мутантных мышей. Иммунофлуоресцентное окрашивание с использованием нефосфорилированного бета-катенина (NPBC) у мышей P30 демонстрирует ядерную экспрессию у Bmp7 ncko и цитоплазматическую экспрессию у мышей Bmp7 ctrl . (C) qRT-PCR носовой перегородки в возрасте P0, P14 и P30 выявила изменения в генах пути Bmp (MSX1, GREM1, ALK2, ALK6). Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: p < 0,05, **: p < 0,01, ***: p < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител.Масштабная линейка = 50 мкм.
Иммуноокрашивание антагонистов Wnt (FRZB, DKK1) у мышей P30 Bmp7 ctrl и Bmp7 ncko демонстрирует увеличение экспрессии FRZB и снижение экспрессии DKK1 у мутантных мышей. Иммунофлуоресцентное окрашивание с использованием нефосфорилированного бета-катенина (NPBC) у мышей P30 демонстрирует ядерную экспрессию у Bmp7 ncko и цитоплазматическую экспрессию у мышей Bmp7 ctrl . (C) qRT-PCR носовой перегородки в возрасте P0, P14 и P30 выявила изменения в генах пути Bmp (MSX1, GREM1, ALK2, ALK6). Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: p < 0,05, **: p < 0,01, ***: p < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител.Масштабная линейка = 50 мкм.
Таким образом, развитие NSD у мышей BMP7 ncko сопровождается переключением в сторону гликолиза наряду с динамическими изменениями сигнальных путей WNT и BMP.
Потеря BMP7 влияет на дифференцировку клеток и клеточный метаболизм уже в носовой перегородке P0
Вышеупомянутые эксперименты показали, что наиболее значительные изменения в экспрессии генов можно наблюдать на P14. Однако некоторые изменения были очевидны уже при рождении. Чтобы систематически идентифицировать изменения экспрессии генов на P0, секвенирование РНК проводили на рассеченных носовых перегородках P0 BMP7 ctrl и BMP7 ncko ( n = 3/генотип).График Volcano идентифицировал как гены с повышенной, так и с отрицательной регуляцией, в то время как биологический коэффициент вариации в образцах составлял от 0,4 до 0,5 в пределах приемлемого диапазона вариаций (рис. 6А). Была создана тепловая карта после неконтролируемой иерархической кластеризации (рис. 6B), а генная онтология (GO) использовалась для сопоставления дифференциально экспрессируемых генов с клеточными процессами, такими как клеточный метаболизм, дифференцировка клеток и морфогенез клеток и структур (рис.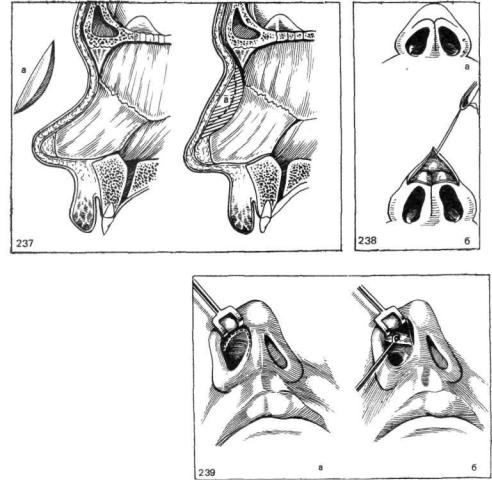 6C). Проверка дифференциально экспрессируемых генов, идентифицированных с помощью анализа RNA-Seq, продемонстрировала изменения пролиферации, апоптоза, клеточного метаболизма, продукции АФК, признаков гипертрофии хондроцитов и передачи сигналов BMP.Отдельные гены, проверенные с помощью RT-qPCR, сопоставлены со значительно измененными онтологиями генов, показаны на рисунке 6D.
6C). Проверка дифференциально экспрессируемых генов, идентифицированных с помощью анализа RNA-Seq, продемонстрировала изменения пролиферации, апоптоза, клеточного метаболизма, продукции АФК, признаков гипертрофии хондроцитов и передачи сигналов BMP.Отдельные гены, проверенные с помощью RT-qPCR, сопоставлены со значительно измененными онтологиями генов, показаны на рисунке 6D.
Рисунок 6. Анализ секвенирования РНК мышей P0 Bmp7 ctrl и Bmp7 ncko , демонстрирующих дифференциальную экспрессию генов ( n = 3/генотип). (A) График вулкана, демонстрирующий гены, по-разному активированные и подавленные. Кроме того, график биологического коэффициента вариации, указывающий, что биологическая изменчивость в образцах находится в пределах от 0.4 и 0,5. (B) Тепловая карта с указанием дифференциально экспрессируемых генов между мышами Bmp7 ctrl и Bmp7 ncko . (C) Генная онтология (GO) — термины, обозначающие гены, идентифицированные как дифференциально экспрессируемые. (D) Определенные термины GO и связанные с ними гены. Измененный метаболизм был важным термином GO, идентифицированным в анализе RNASeq. Гены, выделенные красным, были подтверждены с помощью qRT-PCR (рис. 7).
(D) Определенные термины GO и связанные с ними гены. Измененный метаболизм был важным термином GO, идентифицированным в анализе RNASeq. Гены, выделенные красным, были подтверждены с помощью qRT-PCR (рис. 7).
Ранние признаки измененной клеточной пролиферации, апоптоза, метаболизма и гипертрофии хондроцитов у BMP7
ncko Мыши Сначала мы подтвердили изменения пролиферации и апоптоза в септах P0 у мышей BMP7 ctrl и BMP7 ncko ( n = 3/генотип).Гетерогенный ядерный рибонуклеопротеин I (PTBP1) (Cheung et al., 2009), гетерогенный ядерный рибонуклеопротеин RALY (RALY) (Cornella et al., 2017) и эукариотический фактор инициации трансляции 5A (EIF5A) (Nishimura et al., 2005), известные как стимулирующие пролиферацию клеток, были значительно снижены у мышей BMP7 ncko ( p <0,01, p <0,001, p <0,05) (рис. 7А). Окрашивание на PCNA подтвердило снижение пролиферации хондроцитов.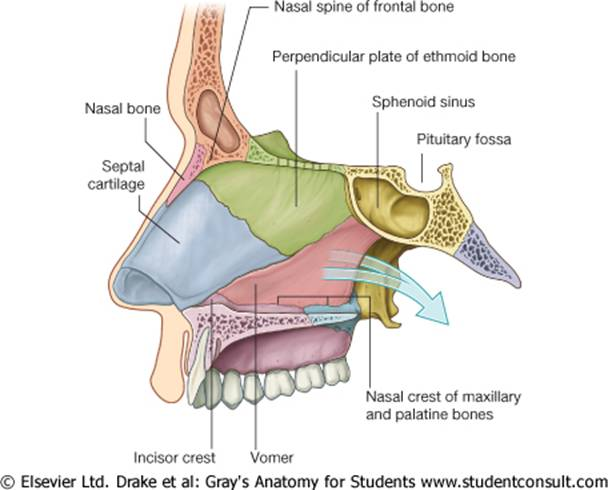 Параллельно экспрессия генов, способствующих выживанию клеток, таких как Component Of Inhibitor Of Nuclear Factor Kinase Kappa B (CHUK) (Culley et al., 2019), TANK Binding Kinase 1 (TBK1) (Xu et al., 2018) и Upstream Фактор транскрипции 1 (USF1) (Guo et al., 2018) также был значительно снижен ( p <0,05), за исключением CHUK (рис. 7B). Это отражалось повышенным уровнем расщепленной каспазы-3 (CASP3) и снижением mTOR (Guo et al., 2018).
Параллельно экспрессия генов, способствующих выживанию клеток, таких как Component Of Inhibitor Of Nuclear Factor Kinase Kappa B (CHUK) (Culley et al., 2019), TANK Binding Kinase 1 (TBK1) (Xu et al., 2018) и Upstream Фактор транскрипции 1 (USF1) (Guo et al., 2018) также был значительно снижен ( p <0,05), за исключением CHUK (рис. 7B). Это отражалось повышенным уровнем расщепленной каспазы-3 (CASP3) и снижением mTOR (Guo et al., 2018).
Рис. 7. Измененная пролиферация клеток, апоптоз и передача сигналов Bmp предшествуют искривлению носовой перегородки. Валидация qRT-PCR на рассеченной носовой перегородке P0 генов, идентифицированных в анализе RNASeq и иммунофлуоресцентном окрашивании, выявила изменение пролиферации клеток (A) , о чем свидетельствует снижение PTBP1, RALY и EIF5a, а также экспрессия PCNA в Bmp7 ncko мышей. (B) Наблюдали повышенный клеточный апоптоз, о чем свидетельствует снижение экспрессии CHUK, TBK1, USF1, повышение экспрессии CASP3 и снижение экспрессии mTOR. (C) Изменения метаболизма липидов наблюдались в виде значительного снижения PRMT1, CHTOP, LIPE и GPD2 с увеличением экспрессии CD44. (D) Наблюдали снижение экспрессии PRDX1 и увеличение экспрессии стрессового белка HSP47. (E) Увеличение MMP13, RUNX2 и COL 1 наблюдалось у мышей Bmp7 ncko . (F) Передача сигналов Bmp, оцененная с помощью иммунофлуоресцентного окрашивания PSMAD, выявила снижение pSMAD у мышей P0 Bmp7 ncko .PTBP1, белок 1, связывающий полипиримидиновый тракт; RALY, гетерогенный ядерный рибонуклеопротеин; EIF5A, эукариотический фактор инициации трансляции 5A; CHUK, компонент комплекса ингибиторов ядерного фактора каппа-киназы B; TBK1, TANK, связывающая киназу 1; USF1, восходящий транскрипционный фактор 1; PRMT1, протеинаргининметилтрансфераза 1; CHTOP, хроматиновая мишень PRMT1; GLI1, цинковый палец семейства GLI 1; LIPE, липаза Е; GPD2, глицерол-3-фосфатдегидрогеназа 2; PRDX1, пероксиредоксин 1; MMP13, матриксная металлопротеиназа 13; RUNX2, транскрипционный фактор семейства RUNX 2; COL 1, коллаген I; PCNA, ядерный антиген пролиферирующих клеток; CASP3, расщепленная каспаза 3; MTOR, мишень рапамицина для млекопитающих; HSP47, СЕРПИНh2; pSMAD, фосфорилированный Smad.
(C) Изменения метаболизма липидов наблюдались в виде значительного снижения PRMT1, CHTOP, LIPE и GPD2 с увеличением экспрессии CD44. (D) Наблюдали снижение экспрессии PRDX1 и увеличение экспрессии стрессового белка HSP47. (E) Увеличение MMP13, RUNX2 и COL 1 наблюдалось у мышей Bmp7 ncko . (F) Передача сигналов Bmp, оцененная с помощью иммунофлуоресцентного окрашивания PSMAD, выявила снижение pSMAD у мышей P0 Bmp7 ncko .PTBP1, белок 1, связывающий полипиримидиновый тракт; RALY, гетерогенный ядерный рибонуклеопротеин; EIF5A, эукариотический фактор инициации трансляции 5A; CHUK, компонент комплекса ингибиторов ядерного фактора каппа-киназы B; TBK1, TANK, связывающая киназу 1; USF1, восходящий транскрипционный фактор 1; PRMT1, протеинаргининметилтрансфераза 1; CHTOP, хроматиновая мишень PRMT1; GLI1, цинковый палец семейства GLI 1; LIPE, липаза Е; GPD2, глицерол-3-фосфатдегидрогеназа 2; PRDX1, пероксиредоксин 1; MMP13, матриксная металлопротеиназа 13; RUNX2, транскрипционный фактор семейства RUNX 2; COL 1, коллаген I; PCNA, ядерный антиген пролиферирующих клеток; CASP3, расщепленная каспаза 3; MTOR, мишень рапамицина для млекопитающих; HSP47, СЕРПИНh2; pSMAD, фосфорилированный Smad. Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: p < 0,05, **: p < 0,01, ***: p < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител. Масштабная линейка = 50 мкм.
Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: p < 0,05, **: p < 0,01, ***: p < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител. Масштабная линейка = 50 мкм.
Protein Arginine Methyltransferase 1 (PRMT1), Chromatin Target Of PRMT1 (CHTOP) и GLI Family Zinc Finger 1 (GLI1) влияют на метаболизм глюкозы (Iwasaki and Yada, 2007; vanLieshout and Ljubicic, 2019).И PRMT1, и CHTOP были значительно снижены ( p <0,001), что свидетельствует о нарушении метаболизма глюкозы уже на P0. И наоборот, гены, участвующие в метаболизме липидов и производстве триглицеридов и холестерина (CD44 (Jiang et al., 2020), липаза E (LIPE) (Gu et al., 2019), митохондриальная глицерол-3-фосфатдегидрогеназа (GPD2) (Mráček et al. ., 2013) также были изменены. В то время как CD44 был значительно повышен ( p < 0,001), LIPE и GPD2 были значительно снижены ( p < 0.01) в мутантных перегородках (рис. 7С). Измененный клеточный метаболизм в хондроцитах был связан с продукцией АФК и реакцией на стресс (Gibson et al., 2008). Действительно, пероксиредоксин-1 (PRDX1), поглотитель АФК (Kumar et al., 2018), был значительно снижен ( p <0,001). Белок теплового шока 47 (HSP47), стрессовый белок, связывающий эндоплазматический коллаген, был повышен в септах BMP7 ncko (рис. 7D). АФК могут вызывать гипертрофию хондроцитов (Morita et al., 2007) в зависимости от BMP (Kumar et al., 2018). Несколько показателей гипертрофии хондроцитов действительно были повышены [матриксная металлопротеиназа-13 (MMP13) ( p <0,001), RUNX2 ( p <0,01), коллаген I (COL 1) ( p <0,001)] (рис. 7E). ), в то время как фосфорилированный SMAD1/5/8 (pSMAD1/5/8) в целом был снижен (рис. 7F). В совокупности это устанавливает, что потеря BMP7 изменяет траекторию развития назальных хондроцитов уже при рождении, задолго до того, как проявляется искривление перегородки.
В то время как CD44 был значительно повышен ( p < 0,001), LIPE и GPD2 были значительно снижены ( p < 0.01) в мутантных перегородках (рис. 7С). Измененный клеточный метаболизм в хондроцитах был связан с продукцией АФК и реакцией на стресс (Gibson et al., 2008). Действительно, пероксиредоксин-1 (PRDX1), поглотитель АФК (Kumar et al., 2018), был значительно снижен ( p <0,001). Белок теплового шока 47 (HSP47), стрессовый белок, связывающий эндоплазматический коллаген, был повышен в септах BMP7 ncko (рис. 7D). АФК могут вызывать гипертрофию хондроцитов (Morita et al., 2007) в зависимости от BMP (Kumar et al., 2018). Несколько показателей гипертрофии хондроцитов действительно были повышены [матриксная металлопротеиназа-13 (MMP13) ( p <0,001), RUNX2 ( p <0,01), коллаген I (COL 1) ( p <0,001)] (рис. 7E). ), в то время как фосфорилированный SMAD1/5/8 (pSMAD1/5/8) в целом был снижен (рис. 7F). В совокупности это устанавливает, что потеря BMP7 изменяет траекторию развития назальных хондроцитов уже при рождении, задолго до того, как проявляется искривление перегородки.
Восстановление BMP2 в BMP7
ncko Спасение мышей при искривлении носовой перегородки При оценке изменений в сигнальном пути BMP (рис. 5C) мы также проверили изменения в экспрессии BMP2, еще одного BMP, участвующего в развитии хряща (Gamer et al., 2018). Действительно, BMP2 повышался на P0 ( p <0,01), не индуцировался на P14, но оставался значительно повышенным на P30 ( p <0,05) ( n = 3/возраст/генотип) (рис. 8A). Было показано, что BMP2 и BMP7 оказывают противоположное влияние на индукцию гипертрофии хондроцитов (Caron et al., 2013). Чтобы оценить, способствует ли увеличение BMP2 непосредственно развитию NSD, мы генетически уменьшили BMP2 у мышей BMP7 ncko (впоследствии BMP2 het BMP7 ncko ).К нашему удивлению, носовая перегородка у мышей P30 BMP2 het BMP7 ncko осталась прямой (рис. 8B, C), отклонение, наблюдаемое у мышей BMP7 ncko , оказалось полностью восстановленным <0,01) (рис. 8D). Некоторые ключевые изменения, выявленные в искривленной перегородке, также были полностью или частично восстановлены (рис. 8E-G). ELN больше не индуцировался (рис. 8E), HNRPU и HK1 продемонстрировали частичную реверсию (рис. 8F), в то время как FRZB и NPBC были сопоставимы с мышами BMP7 ctrl (рис. 8G).Эти находки показывают, что чистый баланс передачи сигналов BMP является критическим для дифференцировки хондроцитов и что этот баланс также влияет на каноническую передачу сигналов WNT, ключевой регулятор энергетического метаболизма, включая гликолиз.
8D). Некоторые ключевые изменения, выявленные в искривленной перегородке, также были полностью или частично восстановлены (рис. 8E-G). ELN больше не индуцировался (рис. 8E), HNRPU и HK1 продемонстрировали частичную реверсию (рис. 8F), в то время как FRZB и NPBC были сопоставимы с мышами BMP7 ctrl (рис. 8G).Эти находки показывают, что чистый баланс передачи сигналов BMP является критическим для дифференцировки хондроцитов и что этот баланс также влияет на каноническую передачу сигналов WNT, ключевой регулятор энергетического метаболизма, включая гликолиз.
Рисунок 8. Снижение BMP2 у мышей Bmp7 ncko устраняет искривление носовой перегородки. (A) qRT-PCR рассеченной носовой перегородки P0, P14, P30 выявил увеличение экспрессии BMP2 на P30 у мышей Bmp7 ncko . (B) Трехмерный вид спереди мыши P30 Bmp2 het Bmp7 ncko , демонстрирующий прямую носовую перегородку. (C) Двухмерный обзор носовой перегородки (обведен белым). (d) Количественная оценка отклонения носового перегородки в BMP2 Het BMP7 NCKO мышей не выявлена существенной разницы между BMP7 Ctrl и BMP2 BMP7 NCKO .Обратите внимание, что данные для Bmp7 ctrl и Bmp7 ncko повторяются с рисунка 1L. (E) Экспрессия эластина (ELN) не наблюдалась у мышей P30 Bmp2 het Bmp7 ncko . (f) Иммунофлуоресцентный окрашивание гетерогенного ядерного рибонуклеопротеина u (HNRPU) в P30 BMP7 CTRL и BMP2 NCKO мышей было сопоставимо, однако, окрашиваясь с гекспокиназой (HK1) снижение экспрессии HK1 у мышей Bmp2 het Bmp7 ncko . (G) Эксплуатация FrzB между P30 BMP7 CTRL и BMP2
(C) Двухмерный обзор носовой перегородки (обведен белым). (d) Количественная оценка отклонения носового перегородки в BMP2 Het BMP7 NCKO мышей не выявлена существенной разницы между BMP7 Ctrl и BMP2 BMP7 NCKO .Обратите внимание, что данные для Bmp7 ctrl и Bmp7 ncko повторяются с рисунка 1L. (E) Экспрессия эластина (ELN) не наблюдалась у мышей P30 Bmp2 het Bmp7 ncko . (f) Иммунофлуоресцентный окрашивание гетерогенного ядерного рибонуклеопротеина u (HNRPU) в P30 BMP7 CTRL и BMP2 NCKO мышей было сопоставимо, однако, окрашиваясь с гекспокиназой (HK1) снижение экспрессии HK1 у мышей Bmp2 het Bmp7 ncko . (G) Эксплуатация FrzB между P30 BMP7 CTRL и BMP2
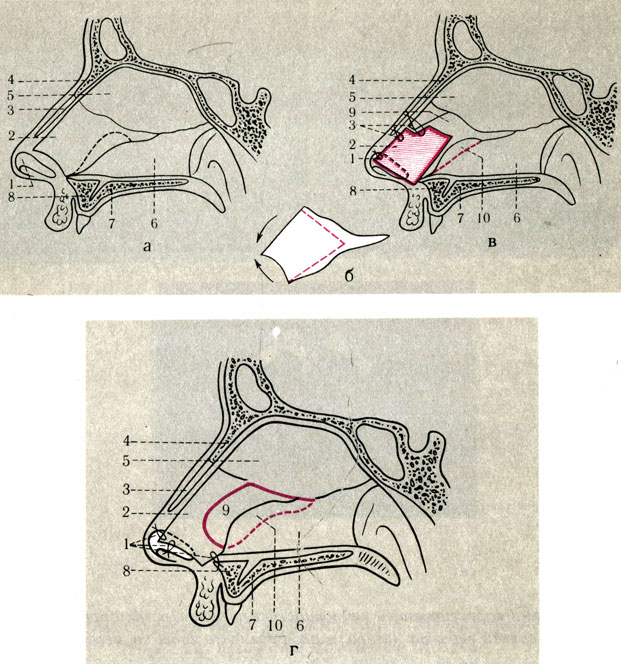 Однако как Bmp7 ctrl , так и Bmp2 het Bmp7 ncko мышей демонстрировали цитоплазматическую экспрессию NPBC.Ins, внутренний шов; ns, носовая перегородка; я, резец; м, нижняя челюсть. Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: P < 0,05, **: P < 0,01, ***: P < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител. Масштабная линейка = 50 мкм.
Однако как Bmp7 ctrl , так и Bmp2 het Bmp7 ncko мышей демонстрировали цитоплазматическую экспрессию NPBC.Ins, внутренний шов; ns, носовая перегородка; я, резец; м, нижняя челюсть. Был проведен независимый двусторонний t -критерий со значимостью, обозначенной как *: P < 0,05, **: P < 0,01, ***: P < 0,001. ( n = 3/генотип/возраст). Экспрессия антител продемонстрирована в красных ядрах, а ядра, окрашенные DAPI, в сером цвете для всех иммунофлуоресцентных изображений. Стрелки обозначают положительный сигнал антител. Масштабная линейка = 50 мкм. Вместе наши данные устанавливают неожиданную сложную этиологию развития NSD, включающую передачу сигналов BMP и WNT.Изменения в этих путях связаны с изменениями в спецификации хондроцитов, последующей дифференцировкой с участием многих клеточных путей, включая клеточный метаболизм, как показано на рисунке 9. Таким образом, при разработке эффективных методов лечения и тканевой инженерии требуется подробная оценка внеклеточного матрикса, клеточного метаболизма и сигнальных свойств.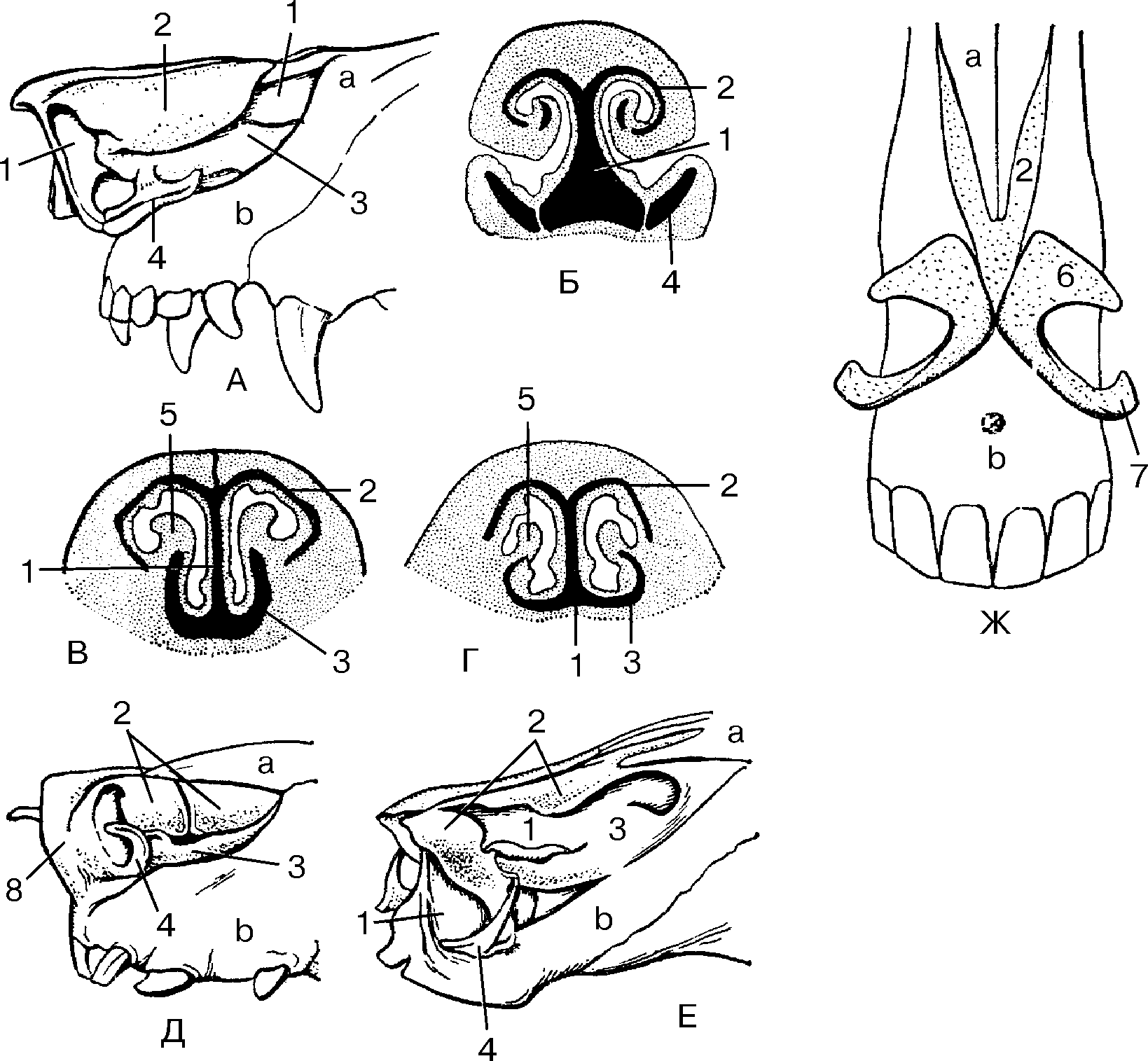 подходы к септопластике.
подходы к септопластике.
Рисунок 9. Изменения хряща, наблюдаемые у мышей Bmp7 ncko . (A) Изменение свойств хряща, передачи сигналов и клеточных процессов наблюдается с течением времени у мышей Bmp7 ncko . Протеомный анализ выявил белки, участвующие в передаче сигналов ретиноевой кислоты через 2 недели (B) и в процессе метаболизма глюкозы через 4 недели (C) .
Обсуждение
В этом исследовании представлена первая подробная характеристика молекулярной и клеточной этиологии НСД в хорошо охарактеризованной модели гипоплазии средней части лица, связанной с обструкцией носовых дыхательных путей (Baddam et al., 2021а). Мы обнаружили, что искривленный хрящ экспрессирует несколько маркеров, в изобилии присутствующих в эластичном хряще, и показал признаки, обычно связанные с остеоартритным хрящом. Это предполагает фундаментальное изменение свойств хряща и подчеркивает сложную этиологию НСД и, вероятно, имеет разветвления для других патологий, связанных с гиалиновым хрящом.
Искривление носовой перегородки классифицируется на основе типа изменения формы (Teixeira et al., 2016; Taghiloo and Halimi, 2019). Некоторые формы НСД возникают в результате нескоординированного роста перегородки и ее сочленяющихся структур (сошника, решетчатой кости, носовых костей), что часто приводит к лицевой асимметрии.В то время как в этих случаях основные дефекты хряща маловероятны, дефекты хряща были предложены в других ситуациях (Teixeira et al., 2016). Остается неясным, почему в некоторых случаях септальный гиалиновый хрящ катастрофически выходит из строя. У мышей BMP7 ncko изначально прямая перегородка уступает место С-образному излому, который дополнительно связан с дефектом роста, приводящим к гипоплазии средней части лица (Baddam et al., 2021a). У новорожденных обычно наблюдаются С- или S-образные отклонения.Неясно, в какой степени аномалии роста хряща уже существуют внутриутробно или могут быть связаны с травмой лица во время рождения (Harugop et al.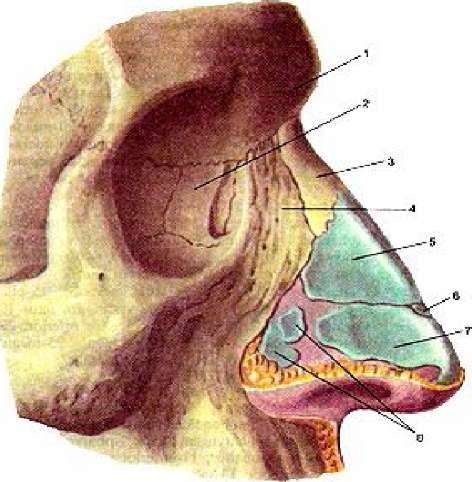 , 2012).
, 2012).
Рост носового хряща у мышей происходит в две фазы. Быстрый рост между P0-P14 и начиная с P21 (Hall and Precious, 2013; Baddam et al., 2021b) прерывается периодом ограниченного роста между ними. Аналогичный бимодальный рост носа наблюдается и у людей, при этом значительный рост происходит в возрасте от 3–4 до 11–12 лет (Albert et al., 2019). Возникновение искривления перегородки у мышей BMP7 ncko до второй фазы роста совпадает с повышенной распространенностью NSD у подростков (Baddam et al., 2020). В то время как первая фаза роста перегородки регулируется пролиферацией клеток, рост во время второй фазы происходит преимущественно за счет гипертрофии клеток (Baddam et al., 2021b). BMP7 экспрессируется в носовом хряще и вокруг него на всех стадиях, с самой низкой экспрессией, наблюдаемой во временной точке P14.Учитывая эту клеточную и молекулярную неоднородность, мы исследовали три временные точки (P0, P14, P30), чтобы понять, проявляются ли и когда изменения носового хряща. Дискретные изменения уже наблюдались при рождении, когда хондроциты были еще незрелыми (Gómez-Picos and Eames, 2015; Baddam et al., 2021b), предполагая, что BMP7 контролирует аспекты ранней дифференцировки хондроцитов, хотя формирующийся хрящ гистологически незаметен, по крайней мере, до P14. Наибольшие различия в экспрессии протеома и генов наблюдались на P14.Значительный апоптоз сопровождался снижением уровня белков маркеров, способствующих гипертрофии хондроцитов (RUNX2 и IHH), и увеличением присутствия белков, ингибирующих гипертрофию (BAPX, FRZB, GREM1) у мышей BMP7 ncko . Эта картина изменилась на П30, где наблюдалось увеличение белков, свидетельствующих о гипертрофии хондроцитов (COLX и IHH). Таким образом, самые ранние идентифицированные изменения уже предполагают изменение свойств хондроцитов в сторону фенотипа эластичного хряща.Около P14, когда рост перегородки переключается с пролиферации на гипертрофический рост, это изменение приводит к катастрофическому апоптозу, гипоплазии средней части лица и из-за измененных свойств хондроцитов к искривлению перегородки.
Дискретные изменения уже наблюдались при рождении, когда хондроциты были еще незрелыми (Gómez-Picos and Eames, 2015; Baddam et al., 2021b), предполагая, что BMP7 контролирует аспекты ранней дифференцировки хондроцитов, хотя формирующийся хрящ гистологически незаметен, по крайней мере, до P14. Наибольшие различия в экспрессии протеома и генов наблюдались на P14.Значительный апоптоз сопровождался снижением уровня белков маркеров, способствующих гипертрофии хондроцитов (RUNX2 и IHH), и увеличением присутствия белков, ингибирующих гипертрофию (BAPX, FRZB, GREM1) у мышей BMP7 ncko . Эта картина изменилась на П30, где наблюдалось увеличение белков, свидетельствующих о гипертрофии хондроцитов (COLX и IHH). Таким образом, самые ранние идентифицированные изменения уже предполагают изменение свойств хондроцитов в сторону фенотипа эластичного хряща.Около P14, когда рост перегородки переключается с пролиферации на гипертрофический рост, это изменение приводит к катастрофическому апоптозу, гипоплазии средней части лица и из-за измененных свойств хондроцитов к искривлению перегородки.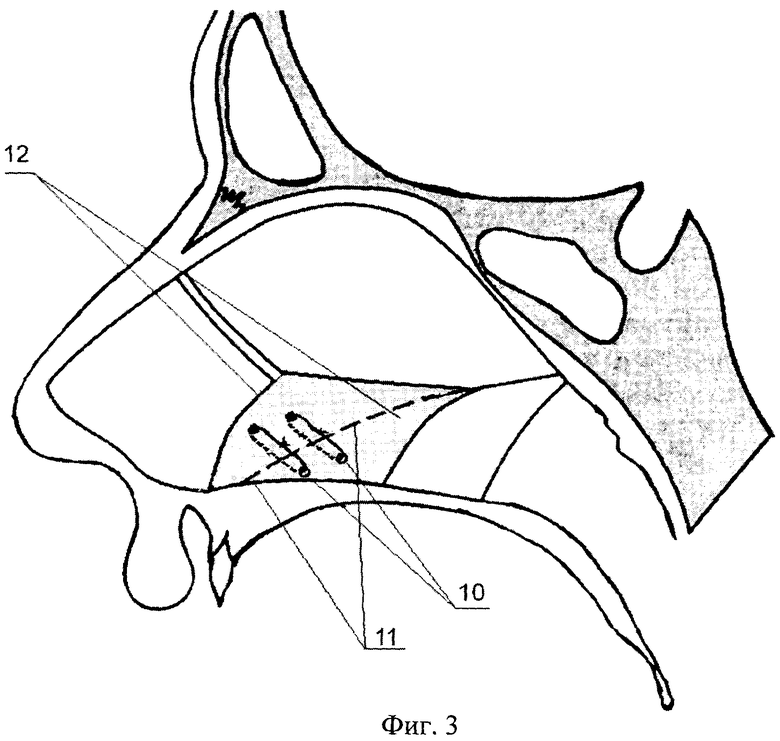 Наши данные совместимы с преобразованием гиалинового хряща в эластичный хрящ и последующей потерей механической устойчивости. Уменьшение сети коллагеновых фибрилл на отклоненной стороне свидетельствует о снижении жесткости (Wong et al., 2018). Контралатеральное увеличение коллагеновых фибрилл может указывать на попытку стабилизации перегородки.Интересно, что и BMP7, и GREM1 (Yu et al., 2017) экспрессируются в местах экспрессии Elastin, что указывает на то, что эти гены могут вносить вклад в ремоделирование хондроцитов и спецификацию хрящей. Большинство изменений первоначально наблюдалось в надхрящнице, а не в зрелых хондроцитах, месте, где находятся клетки-предшественники хряща (Kaucka et al., 2017). Гибель клеток внутри хряща может активировать эти клетки-предшественники хондроцитов для замены умирающих клеток (Seol et al., 2012). Эти перихондральные клетки экспрессируют MFAP5, еще один ген, обнаруженный в эластичном хряще (Asnaghi et al., 2020). Мы предполагаем, что эти экспрессирующие MFAP5 клетки в надхрящнице ускоряют предполагаемый переход от гиалинового к эластичному хрящу.
Наши данные совместимы с преобразованием гиалинового хряща в эластичный хрящ и последующей потерей механической устойчивости. Уменьшение сети коллагеновых фибрилл на отклоненной стороне свидетельствует о снижении жесткости (Wong et al., 2018). Контралатеральное увеличение коллагеновых фибрилл может указывать на попытку стабилизации перегородки.Интересно, что и BMP7, и GREM1 (Yu et al., 2017) экспрессируются в местах экспрессии Elastin, что указывает на то, что эти гены могут вносить вклад в ремоделирование хондроцитов и спецификацию хрящей. Большинство изменений первоначально наблюдалось в надхрящнице, а не в зрелых хондроцитах, месте, где находятся клетки-предшественники хряща (Kaucka et al., 2017). Гибель клеток внутри хряща может активировать эти клетки-предшественники хондроцитов для замены умирающих клеток (Seol et al., 2012). Эти перихондральные клетки экспрессируют MFAP5, еще один ген, обнаруженный в эластичном хряще (Asnaghi et al., 2020). Мы предполагаем, что эти экспрессирующие MFAP5 клетки в надхрящнице ускоряют предполагаемый переход от гиалинового к эластичному хрящу. Потребуется более детальное отслеживание клонов клеток, происходящих из надхрящницы, для дальнейшего понимания механизма, лежащего в основе переключения хряща.
Потребуется более детальное отслеживание клонов клеток, происходящих из надхрящницы, для дальнейшего понимания механизма, лежащего в основе переключения хряща.
Мы использовали количественную протеомику дробовика, чтобы охарактеризовать клеточные последствия этого изменения свойств хряща и понять, почему хондроциты могут подвергаться апоптозу. Сосредоточившись на клеточном метаболизме, мы обнаружили, что mTOR, установленный супрессор аутофагии (Chen and Long, 2018), снижается на P14.У некоторых членов семейства цитохромов P450 (CYP) (Yoshinari et al., 2004; Mao and Zhang, 2018), которые участвуют в поддержании белой жировой ткани, и других белков, участвующих в передаче сигналов ретиноевой кислоты и метаболизме липидов, было увеличено количество белков. BMP7 играет установленную роль в детерминации линии адипоцитов, способствуя коричневому адипогенезу за счет белых адипоцитов (Schulz and Tseng, 2009). Эти две линии адипоцитов характеризуются очень разным клеточным метаболизмом, при этом коричневые адипоциты специализируются на термогенезе за счет разобщения производства митохондриальной энергии. Таким образом, возможно, что BMP7 в назальных хондроцитах сходным образом контролирует спецификацию различных типов хрящей.
Таким образом, возможно, что BMP7 в назальных хондроцитах сходным образом контролирует спецификацию различных типов хрящей.
Описано, что усиление аутофагии и метаболических изменений замедляет прогрессирование ОА (Luo et al., 2019). Таким образом, эти два изменения могут отражать защитные механизмы, замедляющие деградацию хряща в перегородке. Неясно, проявляются ли эти изменения во всех или только в подмножестве хондроцитов, происходят ли эти изменения в одном и том же хондроците и связаны ли эти изменения непосредственно с наблюдаемой гибелью клеток.Подсказка может быть получена из данных P30, где наблюдалось увеличение классических маркеров гипертрофии хондроцитов (IHH, COLX), а также активация белков, участвующих в гликолизе. Hollander and Zeng (2019) продемонстрировали, что гипертрофированные хондроциты экспрессируют более высокие уровни глицеральдегид-3-фосфатдегидрогеназы (GAPDH) и лактатдегидрогеназы (LDH), двух белков с повышенной активностью у мышей P30 BMP7 ncko .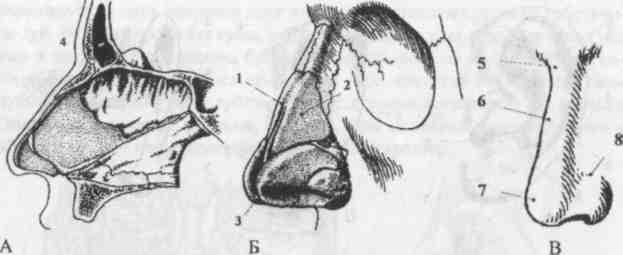 Не исключено, что на Р14 выживают клетки, перешедшие на гликолиз, а клетки с повышенным метаболизмом липидов обречены на гибель.В то время как необходимы дальнейшие исследования, чтобы разрешить эти возможности, наши данные убедительно свидетельствуют о том, что BMP7 контролирует раннюю дифференцировку хондроцитов по разным траекториям развития (гиалиновый против эластичного хряща). Необходимо показать, соответствуют ли эти траектории действительно разным клеточным линиям, как описано для коричневых/белых адипоцитов.
Не исключено, что на Р14 выживают клетки, перешедшие на гликолиз, а клетки с повышенным метаболизмом липидов обречены на гибель.В то время как необходимы дальнейшие исследования, чтобы разрешить эти возможности, наши данные убедительно свидетельствуют о том, что BMP7 контролирует раннюю дифференцировку хондроцитов по разным траекториям развития (гиалиновый против эластичного хряща). Необходимо показать, соответствуют ли эти траектории действительно разным клеточным линиям, как описано для коричневых/белых адипоцитов.
Было показано, что BMP7 не является единственным BMP, контролирующим дифференцировку адипоцитов. BMP2 является индуктором белой жировой ткани (Blázquez-Medela et al., 2019). BMP2 и BMP7 принадлежат к разным подгруппам BMP. В то время как первый является ортологом Drosophila Melanogaster Decapentaplegic, последний является ортологом стеклянного дна и принадлежит к группе 60A (Malik et al., 2020). В контрольной перегородке BMP7 BMP2 динамически экспрессировался во время созревания носовой перегородки, пик экспрессии наблюдался на P14, а экспрессия была низкой на P30.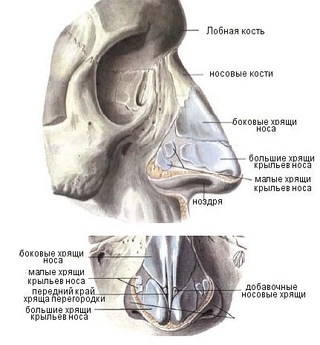 У мышей BMP7 ncko индукция BMP2 на P14 была остановлена, но экспрессия была существенно увеличена на P30.Увеличение BMP2 на P30 в хряще с дефицитом BMP7 может свидетельствовать о попытке молекулярной компенсации. Если это так, то можно ожидать, что снижение BMP2 у мышей BMP7 ncko приведет к более тяжелому НСД. Мыши BMP2/BMP7 с нокаутом двойного нервного гребня умирают при рождении. Таким образом, мы создали гетерозиготных мышей BMP2/BMP7 ncko . Снижение BMP2 предотвращало приобретение маркеров эластичности и частично восстанавливало клеточный метаболизм. Было показано, что в первичных хондроцитах BMP2 увеличивает метаболизм глюкозы во время эндохондральной оссификации (Lee et al., 2018), гипертрофия хондропрогениторных клеток (Caron et al., 2013) и была связана с деградацией хряща при ОА (Mariani et al., 2014). Таким образом, мы предполагаем, что баланс между BMP2 и BMP7 скорее, чем отдельные сигналы, контролирует метаболизм и дифференцировку хондроцитов.
У мышей BMP7 ncko индукция BMP2 на P14 была остановлена, но экспрессия была существенно увеличена на P30.Увеличение BMP2 на P30 в хряще с дефицитом BMP7 может свидетельствовать о попытке молекулярной компенсации. Если это так, то можно ожидать, что снижение BMP2 у мышей BMP7 ncko приведет к более тяжелому НСД. Мыши BMP2/BMP7 с нокаутом двойного нервного гребня умирают при рождении. Таким образом, мы создали гетерозиготных мышей BMP2/BMP7 ncko . Снижение BMP2 предотвращало приобретение маркеров эластичности и частично восстанавливало клеточный метаболизм. Было показано, что в первичных хондроцитах BMP2 увеличивает метаболизм глюкозы во время эндохондральной оссификации (Lee et al., 2018), гипертрофия хондропрогениторных клеток (Caron et al., 2013) и была связана с деградацией хряща при ОА (Mariani et al., 2014). Таким образом, мы предполагаем, что баланс между BMP2 и BMP7 скорее, чем отдельные сигналы, контролирует метаболизм и дифференцировку хондроцитов.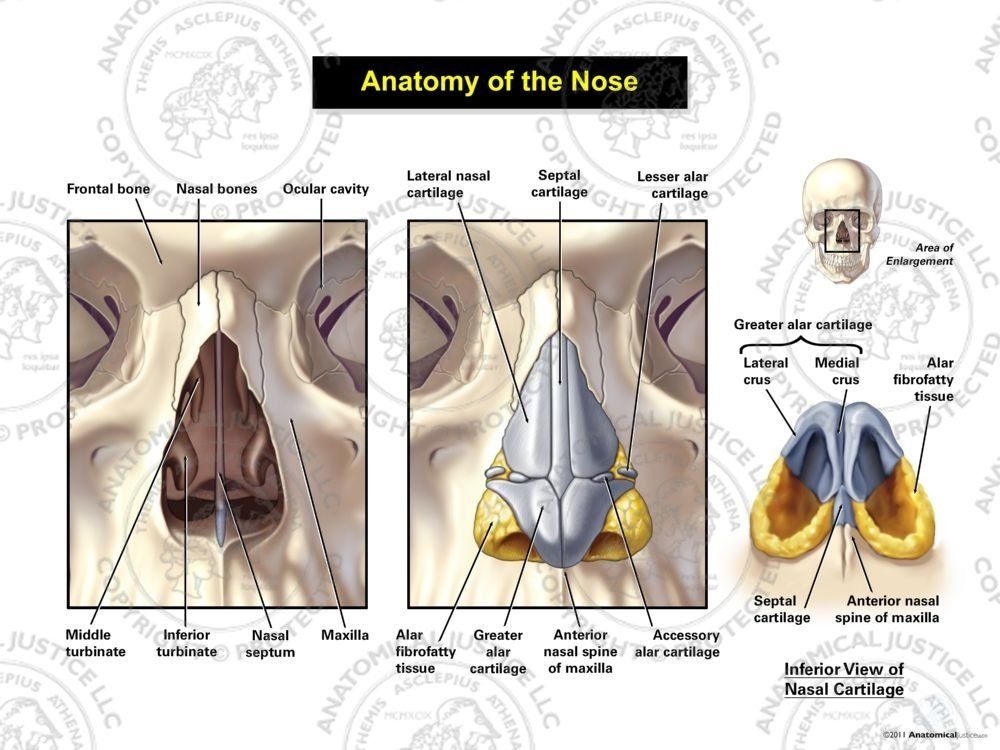 Наши результаты не только иллюстрируют, как метаболические потребности меняются во время созревания хондроцитов, но и сами метаболические различия, по-видимому, связаны с различными свойствами хряща.Является ли измененный метаболизм причиной изменения свойств хондроцитов или является следствием измененных свойств хондроцитов, еще предстоит установить. Гипертрофия хондроцитов (COLX, IHH) и изменение метаболизма глюкозы, наблюдаемое при искривлении перегородки, являются отличительными признаками ОА коленного сустава (van der Kraan and van den Berg, 2012). Кроме того, было замечено несколько других сходств. COL2 и OPN снижены как при ОА коленного сустава (Tsolis et al., 2015; Liu et al., 2020), так и при НСД. COL VI, белок перицеллюлярного матрикса, активность которого повышается при ОА (Alexopoulos et al., 2009) также увеличилась в НРД. Изменения во внеклеточном матриксе могут влиять на механоощущение, приводящее к клеточному стрессу (Tsolis et al., 2015), и могут непосредственно способствовать увеличению апоптоза.
Наши результаты не только иллюстрируют, как метаболические потребности меняются во время созревания хондроцитов, но и сами метаболические различия, по-видимому, связаны с различными свойствами хряща.Является ли измененный метаболизм причиной изменения свойств хондроцитов или является следствием измененных свойств хондроцитов, еще предстоит установить. Гипертрофия хондроцитов (COLX, IHH) и изменение метаболизма глюкозы, наблюдаемое при искривлении перегородки, являются отличительными признаками ОА коленного сустава (van der Kraan and van den Berg, 2012). Кроме того, было замечено несколько других сходств. COL2 и OPN снижены как при ОА коленного сустава (Tsolis et al., 2015; Liu et al., 2020), так и при НСД. COL VI, белок перицеллюлярного матрикса, активность которого повышается при ОА (Alexopoulos et al., 2009) также увеличилась в НРД. Изменения во внеклеточном матриксе могут влиять на механоощущение, приводящее к клеточному стрессу (Tsolis et al., 2015), и могут непосредственно способствовать увеличению апоптоза.
WNT контролирует клеточный рост, дифференцировку, подвижность и клеточный метаболизм. Было показано, что в хряще каноническая передача сигналов WNT способствует гипертрофии хондроцитов (Mariani et al., 2014; Houben et al., 2016; Liu et al., 2016). Поскольку передача сигналов BMP и WNT часто взаимодействует (Choe et al., 2013; Malik et al., 2020), мы исследовали изменения в генах пути WNT. Потеря BMP7 привела к изменению экспрессии нескольких лигандов WNT (увеличение WNT6 и WNT7A, снижение WNT3A), антагонистов WNT (увеличение FRZB, снижение DKK1) и общей активации канонической передачи сигналов WNT, о чем свидетельствует ядерная локализация не- фосфорилированный β-катенин (Choe et al., 2013). Снижение BMP2, помимо предотвращения экспрессии эластина и метаболических изменений, также корректирует каноническую передачу сигналов WNT.Какой из этих эффектов является прямым следствием восстановленной передачи сигналов WNT или является прямым результатом перебалансированной передачи сигналов BMP, в настоящее время неясно. Например, отклонение перегородки связано с повышенной механической нагрузкой (Yang et al., 2017), что само по себе связано с усилением канонической передачи сигналов WNT (Brunt et al., 2017). Тем не менее, спасение NSD подчеркивает необходимость понимания того, как эта сбалансированная передача сигналов контролирует нормальный носовой и, возможно, более широкий рост средней части лица. Выявление общих черт и различий между передачей сигналов BMP и WNT будет иметь решающее значение для лучшего понимания этой молекулярной сложности.Тем не менее, это исследование четко устанавливает критическое участие передачи сигналов BMP и WNT в этиологии NSD. Величина и время изменений в этих сигнальных путях могут определять начало и тяжесть НСД и связанных с ним симптомов.
Например, отклонение перегородки связано с повышенной механической нагрузкой (Yang et al., 2017), что само по себе связано с усилением канонической передачи сигналов WNT (Brunt et al., 2017). Тем не менее, спасение NSD подчеркивает необходимость понимания того, как эта сбалансированная передача сигналов контролирует нормальный носовой и, возможно, более широкий рост средней части лица. Выявление общих черт и различий между передачей сигналов BMP и WNT будет иметь решающее значение для лучшего понимания этой молекулярной сложности.Тем не менее, это исследование четко устанавливает критическое участие передачи сигналов BMP и WNT в этиологии NSD. Величина и время изменений в этих сигнальных путях могут определять начало и тяжесть НСД и связанных с ним симптомов.
Таким образом, наши результаты показывают, что многофакторная этиология лежит в основе НСД, связанной с гипоплазией средней части лица. В настоящее время неясно, какие из этих изменений являются прямым следствием отсутствия BMP7, а какие развиваются как следствие. Также неясно, предрасполагают ли клеточные изменения (например, клеточный метаболизм, гипертрофия) или связанные с ними изменения на уровне ткани (например, более тонкая перегородка) или приводят к искривлению перегородки. Модель мыши BMP7 ncko представляет собой генетическую основу, имеющую отношение к развитию хряща и патологиям, касающимся не только носовой перегородки, но и гиалинового хряща в целом. Это также указывает на возможность того, что тип хряща определяется на ранней стадии развития и что BMP7 может быть фактором, способствующим этому.Наши данные также свидетельствуют о том, что хрящи различного происхождения могут иметь сходство, но по-разному запрограммированы для удовлетворения требований места, в котором находится хрящ. суставного хряща для лечения ОА. Отныне изменения, описанные в мышиной модели BMP7 ncko до и после установления отклонения перегородки, могут иметь отношение к лучшему пониманию ОА и других патологий хряща.Были достигнуты значительные успехи в понимании эндохондральной оссификации и трансдифференцировки хряща в кость; однако до сих пор неясно, как разные типы хрящей специфицируются из предшественников хондроцитов.
Также неясно, предрасполагают ли клеточные изменения (например, клеточный метаболизм, гипертрофия) или связанные с ними изменения на уровне ткани (например, более тонкая перегородка) или приводят к искривлению перегородки. Модель мыши BMP7 ncko представляет собой генетическую основу, имеющую отношение к развитию хряща и патологиям, касающимся не только носовой перегородки, но и гиалинового хряща в целом. Это также указывает на возможность того, что тип хряща определяется на ранней стадии развития и что BMP7 может быть фактором, способствующим этому.Наши данные также свидетельствуют о том, что хрящи различного происхождения могут иметь сходство, но по-разному запрограммированы для удовлетворения требований места, в котором находится хрящ. суставного хряща для лечения ОА. Отныне изменения, описанные в мышиной модели BMP7 ncko до и после установления отклонения перегородки, могут иметь отношение к лучшему пониманию ОА и других патологий хряща.Были достигнуты значительные успехи в понимании эндохондральной оссификации и трансдифференцировки хряща в кость; однако до сих пор неясно, как разные типы хрящей специфицируются из предшественников хондроцитов. Понимание спецификации происхождения хряща облегчит клиническую проблему формирования волокнистого хряща, несмотря на усилия по регенерации гиалинового хряща для лечения ОА (Armiento et al., 2019). Кроме того, стратегии тканевой инженерии на основе клеток, такие как имплантация аутологичных хондроцитов (ACI) (Brittberg et al., 1994; Mumme et al., 2016) или его более поздний вариант, имплантация хондроцитов с матриксом (MACI) (Behrens et al., 2006; Knutsen et al., 2007; Zheng et al., 2007), для восстановления очаговых поражений суставного хряща. значительно выиграет от понимания клеточных процессов, которые одновременно способствуют формированию гиалинового хряща и уменьшают образование волокнистого хряща.
Понимание спецификации происхождения хряща облегчит клиническую проблему формирования волокнистого хряща, несмотря на усилия по регенерации гиалинового хряща для лечения ОА (Armiento et al., 2019). Кроме того, стратегии тканевой инженерии на основе клеток, такие как имплантация аутологичных хондроцитов (ACI) (Brittberg et al., 1994; Mumme et al., 2016) или его более поздний вариант, имплантация хондроцитов с матриксом (MACI) (Behrens et al., 2006; Knutsen et al., 2007; Zheng et al., 2007), для восстановления очаговых поражений суставного хряща. значительно выиграет от понимания клеточных процессов, которые одновременно способствуют формированию гиалинового хряща и уменьшают образование волокнистого хряща.
Заявление о доступности данных
Протеомные данные были переданы в Консорциум ProteomeXchange через партнерский репозиторий PRIDE под номером доступа PXD024813.Для данных RNAseq необработанные данные секвенирования общедоступны на портале NCBI SRA под номером доступа PRJNA622501.
Заявление об этике
Исследование на животных было рассмотрено и одобрено Комитетом медицинских наук по уходу за животными и их использованию Университета Альберты (протокол №: AUP1149).
Вклад авторов
PB: разработка исследований, проведение экспериментов, сбор данных, анализ данных, написание рукописи и редактирование рукописи.DY: проведение экспериментов, сбор данных, анализ данных, написание рукописи и редактирование рукописи. GD: анализ данных и редактирование рукописи. CN: анализ данных и редактирование рукописи. FE: проведение экспериментов и редактирование рукописи. SE: редактирование рукописи. JJ: проведение экспериментов, предоставление реагентов, сбор данных, анализ данных и редактирование рукописи. АА: разработка исследований, анализ данных и редактирование рукописи. AD: анализ данных, предоставление реагентов, редактирование рукописи и получение финансирования.DG: разработка исследований, предоставление реагентов, анализ данных, написание рукописи, редактирование рукописи и получение финансирования.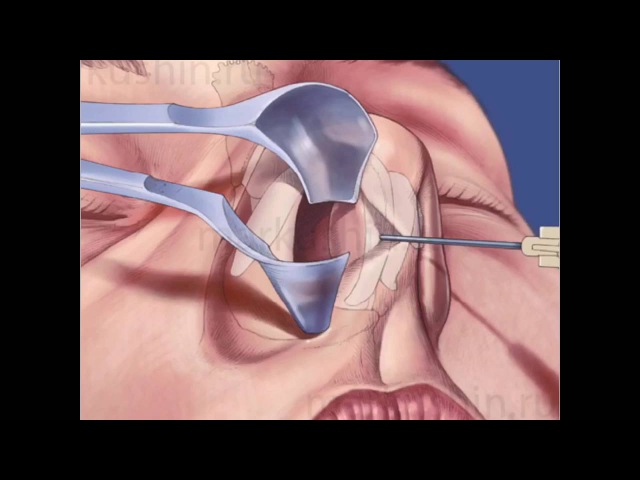 Все авторы внесли свой вклад в статью и одобрили представленную версию.
Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа была поддержана грантом RGPIN-2014-06311 (DG) Совета по естественным наукам и инженерным исследованиям (NSERC), инновационным грантом (DG) Исследовательского института здоровья женщин и детей (WCHRI), исследовательской группой Джеффри и Робин Спербер по черепно-лицевой биологии. Фонды (DG), грант NSERC на открытие CGECR-2019-00112 (AD).Авторы также хотели бы отметить поддержку стажеров в рамках аспирантской стипендии NSERC (PB) и летней студенческой программы NSERC (CN).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы признательны лаборатории мкКТ Школы стоматологии Университета Альберты.Мы благодарим Luc Berthiaume (кафедра клеточной биологии Университета Альберты) за любезно предоставленное антитело против GFP.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fcell.2021.696545/full#supplementary-material
.Сноски
Каталожные номера
Абула К., Мунета Т., Миятаке К., Ямада Дж., Мацукура Ю., Иноуэ М. и др. (2015).Элиминация BMP7 из развивающейся мезенхимы конечностей приводит к дегенерации суставного хряща и синовиальному воспалению с возрастом. ФЭБС Письмо. 589, 1240–1248. doi: 10.1016/j.febslet.2015.04.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Альберт, А. М., Пейн, А. Л., Брэди, С. М., и Райт, К. (2019). Черепно-лицевые изменения у детей от рождения до позднего подросткового возраста. ARC J. Forensic Sci. 4, 1–19.
Академия Google
Алексопулос, Л.Г., Юн И., Бональдо П. и Гуилак Ф. (2009). Изменения развития и остеоартрита у мышей с нокаутом Col6a1: биомеханика коллагена VI типа в перицеллюлярном матриксе хряща. Ревматоидный артрит. 60, 771–779. дои: 10.1002/арт.24293
Ревматоидный артрит. 60, 771–779. дои: 10.1002/арт.24293
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Алсуфьяни, Н., Исаак, А., Витманс, М., Мейджор, П., и Эль-Хаким, Х. (2017). Предикторы неудачи DISE-направленной аденотонзиллэктомии у детей с нарушениями дыхания во сне. Ж. Отоларингол. Хирургия головы и шеи. 46:37. doi: 10.1186/s40463-017-0213-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Армиенто, А. Р., Алини, М., и Стоддарт, М. Дж. (2019). Суставной волокнистый хрящ. Почему гиалиновый хрящ не восстанавливается? Доп. Доставка лекарств, ред. 146, 289–305. doi: 10.1016/j.addr.2018.12.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аснаги, М. А., Пауэр, Л., Барберо, А., Haug, M., Koppl, R., Wendt, D., et al. (2020). Подписи биомаркеров качества для разработки носового хряща, полученного из хондроцитов. Фронт. биоинж. Биотехнолог. 8:283. doi: 10.3389/fbioe.2020.00283
8:283. doi: 10.3389/fbioe.2020.00283
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Баддам, П., Бьянкарди, В., Рот, Д.М., Итон, Ф., Тереза-Буссоларо, К., Мандал, Р., и др. (2021а). Делеция Bmp7, специфичная для нервного гребня, приводит к гипоплазии средней части лица, обструкции носовых дыхательных путей и нарушению дыхания, моделирующему обструктивное апноэ во сне. Дис. Модель мех. 14:dmm047738. doi: 10.1242/dmm.047738
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Баддам, П., Буссоларо, К.-Т., Флорес-Мир, К., и Граф, Д. (2020). Структурные аномалии полости носа у детей с высоким риском нарушений дыхания во сне: исследование с помощью конусно-лучевой компьютерной томографии. утра. Дж. Ортодонт. Зубочелюстной ортопед. [в печати].
Академия Google
Баддам П., Кунг Т., Адесида А.Б. и Граф Д. (2021b). Гистологическая и молекулярная характеристика растущей носовой перегородки у мышей.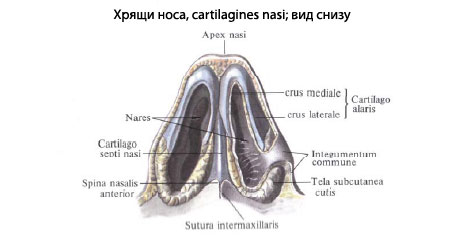 Дж. Анат. 238, 751–764. doi: 10.1111/joa.13332
Дж. Анат. 238, 751–764. doi: 10.1111/joa.13332
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Беренс П., Биттер Т., Курц Б. и Расслис М. (2006). Ассоциированная с матриксом трансплантация/имплантация аутологичных хондроцитов (MACT/MACI) – 5-летнее наблюдение. Колено 13, 194–202. doi: 10.1016/j.knee.2006.02.012
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бласкес-Медела, А.М., Джумабай М. и Бострём К.И. (2019). За пределами кости: передача сигналов костного морфогенетического белка (BMP) в жировой ткани. OBES Ред. 20, 648–658. doi: 10.1111/обр.12822
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бриттберг М., Линдал А., Нильссон А., Олссон С., Исакссон О. и Петерсон Л. (1994). Лечение глубоких дефектов хряща коленного сустава трансплантацией аутологичных хондроцитов. Н. англ. Дж. Мед. 331, 889–895. дои: 10.1056/NEJM199410063311401
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Брант, Л. Х., Бегг, К., Каге, Э., Кросс, С., и Хаммонд, К. Л. (2017). Передача сигналов Wnt контролирует реакцию на механическую нагрузку во время развития суставов у рыбок данио. Разработка 144, 2798–2809. doi: 10.1242/dev.153528
Х., Бегг, К., Каге, Э., Кросс, С., и Хаммонд, К. Л. (2017). Передача сигналов Wnt контролирует реакцию на механическую нагрузку во время развития суставов у рыбок данио. Разработка 144, 2798–2809. doi: 10.1242/dev.153528
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кандела, М.Э., Ван, К., Гунавардена, А.Т., Чжан, К., Кэнтли, Л., Ясухара, Р., и другие. (2016). Интегрин альфа-5 опосредует остеоартритные изменения в коленных суставах мышей. PLoS One 11:e0156783. doi: 10.1371/journal.pone.0156783
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Caron, M.M.J., Emans, P.J., Cremers, A., Surtel, D.A.M., Coolsen, M.M.E., van Rhijn, L.W., et al. (2013). Гипертрофическая дифференцировка во время хондрогенной дифференцировки клеток-предшественников стимулируется BMP-2, но подавляется BMP-7. Остеоартрит.Хрящ 21, 604–613. doi: 10.1016/j.joca.2013.01.009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Карон, М.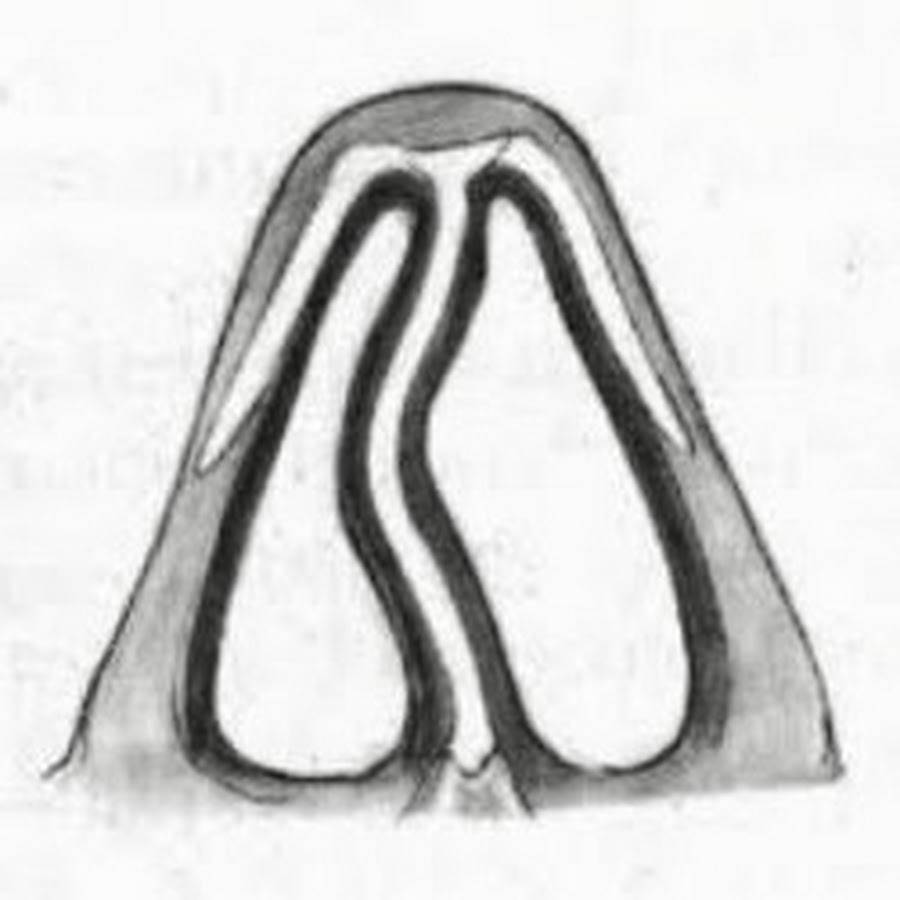 М.Дж., Эманс, П.Дж., Суртел, Д.А.М., ван дер Краан, П.М., ван Рейн, Л.В., и Велтинг, Т.Дж.М. (2015). BAPX-1/NKX-3.2 действует как молекулярный переключатель гипертрофии хондроцитов при остеоартрите. Артрит Ревматолог. 67, 2944–2956. doi: 10.1002/art.39293
М.Дж., Эманс, П.Дж., Суртел, Д.А.М., ван дер Краан, П.М., ван Рейн, Л.В., и Велтинг, Т.Дж.М. (2015). BAPX-1/NKX-3.2 действует как молекулярный переключатель гипертрофии хондроцитов при остеоартрите. Артрит Ревматолог. 67, 2944–2956. doi: 10.1002/art.39293
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чунг, Х.C., Hai, T., Zhu, W., Baggerly, K.A., Tsavachidis, S., Krahe, R., et al. (2009). Факторы сплайсинга PTBP1 и PTBP2 способствуют пролиферации и миграции клеточных линий глиомы. Мозг 132, 2277–2288. дои: 10.1093/мозг/awp153
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чое, Ю., Козлова, А., Граф, Д., и Удовольствие, С. Дж. (2013). Передача сигналов костного морфогенного белка является основной детерминантой развития зубчатых зубов. J. Neurosci. 33, 6766–6775.doi: 10.1523/JNEUROSCI.0128-13.2013
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Коулз, Дж. М., Чжан, Л., Блюм, Дж. Дж., Варман, М. Л., Джей, Г. Д., Гуилак, Ф., и соавт. (2010). Потеря структуры хряща, жесткости и фрикционных свойств у мышей, лишенных PRG4. Ревматоидный артрит. 62, 1666–1674. дои: 10.1002/арт.27436
М., Чжан, Л., Блюм, Дж. Дж., Варман, М. Л., Джей, Г. Д., Гуилак, Ф., и соавт. (2010). Потеря структуры хряща, жесткости и фрикционных свойств у мышей, лишенных PRG4. Ревматоидный артрит. 62, 1666–1674. дои: 10.1002/арт.27436
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Корнелла, Н., Тебальди, Т., Гасперини, Л., Сингх, Дж., Padgett, R.A., Rossi, A., et al. (2017). hnRNP RALY регулирует транскрипцию и пролиферацию клеток, модулируя экспрессию специфических факторов, включая маркер пролиферации E2F1. Дж. Биол. хим. 292, 19674–19692. doi: 10.1074/jbc.M117.795591
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кокс, Дж., и Манн, М. (2008). MaxQuant обеспечивает высокую скорость идентификации пептидов, индивидуальную точность массы в диапазоне частей на миллиард и количественную оценку белков в масштабах всего протеома. Нац. Биотехнолог. 26, 1367–1372. doi: 10.1038/nbt.1511
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кокс, Дж. , Нойхаузер, Н., Михальский, А., Шелтема, Р. А., Олсен, Дж. В., и Манн, М. (2011). Andromeda: поисковая система пептидов, интегрированная в среду MaxQuant. J. Proteome Res. 10, 1794–1805 гг. дои: 10.1021/pr101065j
, Нойхаузер, Н., Михальский, А., Шелтема, Р. А., Олсен, Дж. В., и Манн, М. (2011). Andromeda: поисковая система пептидов, интегрированная в среду MaxQuant. J. Proteome Res. 10, 1794–1805 гг. дои: 10.1021/pr101065j
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Калли, К.Л., Лессард, С.Г., Грин, Дж. Д., Куинн, Дж., Чанг, Дж., Хилнани, Т., и соавт. (2019). Индуцируемый нокаут CHUK/IKKα во взрослых хондроцитах уменьшает прогрессирование деградации хряща в хирургической модели остеоартрита. науч. Респ. 9:8905. doi: 10.1038/s41598-019-45334-5
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Д’Асканио, Л., Лансионе, К., Помпа, Г., Ребуффини, Э., Манси, Н., и Манзини, М. (2010). Черепно-лицевой рост у детей с искривлением носовой перегородки: цефалометрическое сравнительное исследование. Междунар. Дж. Педиатр. Оториноларингол. 74, 1180–1183. doi: 10.1016/j.ijporl.2010.07.010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Геймер, Л. В., Прегизер, С., Геймер, Дж., Фейгенсон, М., Ионеску, А., Ли, К., и соавт. (2018). Роль Bmp2 в созревании и поддержании коленного сустава мыши. J. Bone Miner Res. 33, 1708–1717. doi: 10.1002/jbmr.3441
В., Прегизер, С., Геймер, Дж., Фейгенсон, М., Ионеску, А., Ли, К., и соавт. (2018). Роль Bmp2 в созревании и поддержании коленного сустава мыши. J. Bone Miner Res. 33, 1708–1717. doi: 10.1002/jbmr.3441
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Герстенфельд, Л.C. и Шапиро Ф.Д. (1996). Экспрессия костно-специфических генов гипертрофическими хондроцитами: влияние сложных функций гипертрофических хондроцитов на развитие эндохондральной кости. J. Cell Biochem. 62, 1–9. doi: 10.1002/(SICI)1097-4644(199607)62:1<1::AID-JCB1<3.0.CO;2-X
Полнотекстовая перекрестная ссылка | Академия Google
Гибсон, Дж. С., Милнер, П. И., Уайт, Р., Фэйрфакс, Т. П. А., и Уилкинс, Р. Дж. (2008). Кислород и активные формы кислорода в суставном хряще: модуляторы ионного гомеостаза. Арка Пфлюгера. 455, 563–573. doi: 10.1007/s00424-007-0310-7
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gu, H. -Y., Yang, M., Guo, J., Zhang, C., Lin, L.-L., Liu, Y., et al. (2019). Идентификация биомаркеров и патологического процесса остеоартрита: анализ сети коэкспрессии взвешенных генов. Фронт. Физиол. 10:00275. doi: 10.3389/fphys.2019.00275
-Y., Yang, M., Guo, J., Zhang, C., Lin, L.-L., Liu, Y., et al. (2019). Идентификация биомаркеров и патологического процесса остеоартрита: анализ сети коэкспрессии взвешенных генов. Фронт. Физиол. 10:00275. doi: 10.3389/fphys.2019.00275
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Го, Дж., Fang, W., Chen, X., Lin, Y., Hu, G., Wei, J., et al. (2018). Восходящий стимулирующий фактор 1 подавляет аутофагию и катаболизм капель липидов в печени путем активации mTOR. ФЭБС Письмо. 592, 2725–2738. дои: 10.1002/1873-3468.13203
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холл, Б.К., и Драгоценный, Д.С. (2013). Расщелина губы, носа и неба: носовая перегородка как стимулятор роста средней части лица. Оральный хирург. Оральный мед. Орал Патол. Оральный радиол. 115, 442–447. doi: 10.1016/j.oooo.2012.05.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Харугоп, А. С., Мудхол, Р.С., Хаджаре, П.С., Наргунд, А.И., Метгудмат, В.В., и Чакрабарти, С. (2012). Распространенность искривления носовой перегородки у новорожденных и его провоцирующие факторы: перекрестное исследование. Ind. J. Отоларингол. Хирургия головы и шеи. 64, 248–251. doi: 10.1007/s12070-011-0247-1
С., Мудхол, Р.С., Хаджаре, П.С., Наргунд, А.И., Метгудмат, В.В., и Чакрабарти, С. (2012). Распространенность искривления носовой перегородки у новорожденных и его провоцирующие факторы: перекрестное исследование. Ind. J. Отоларингол. Хирургия головы и шеи. 64, 248–251. doi: 10.1007/s12070-011-0247-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хубен, А., Костанова-Полякова Д., Вайссенбёк М., Граф Дж., Тойфель С., Марк К. и соавт. (2016). Активность β-катенина в поздних гипертрофированных хондроцитах локально управляет остеобластогенезом и остеокластогенезом. Разработка 143, 3826–3838. doi: 10.1242/dev.137489
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хуанг, X., Чжун, Л., Пост, Дж. Н., и Карпериен, М. (2018). Совместное лечение TGF-β3 и BMP7 лучше стимулирует редифференцировку хондроцитов как при гипоксии, так и при нормоксии по сравнению с однократным лечением. науч. Респ. 8:10251.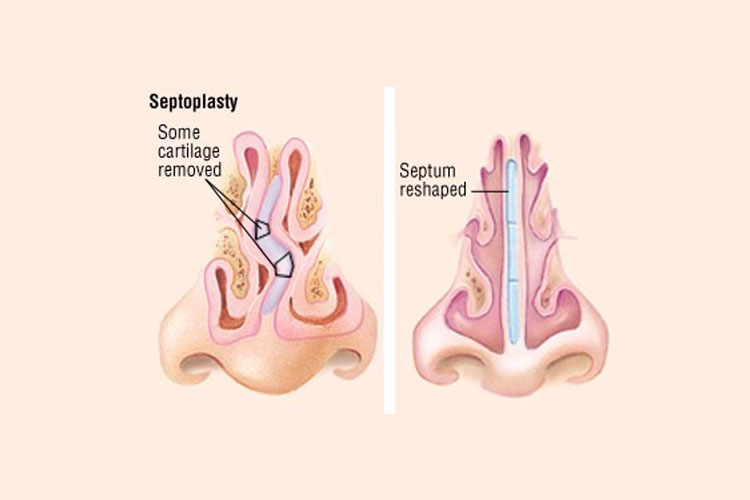 doi: 10.1038/s41598-018-27602-y
doi: 10.1038/s41598-018-27602-y
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Исикава Ю., Рубин К., Бехингер Х. П. и Каламайски С. (2018). Находящийся в эндоплазматическом ретикулуме коллагеновый шаперон Hsp47 взаимодействует и способствует секреции декорина, фибромодулина и люмикана. Дж. Биол. хим. 293, 13707–13716. doi: 10.1074/jbc.RA117.000758
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ивасаки, Х.и Яда, Т. (2007). Метилирование аргинина в белке регулирует передачу сигналов инсулина в клетках скелетных мышц L6. Биохим. Биофиз. Рез. коммун. 364, 1015–1021. doi: 10.1016/j.bbrc.2007.10.113
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Цзян П., Ся Л., Джин З., Али С., Ван М., Ли Х. и др. (2020). Новая функция гена CD44: регуляция липидного обмена в эпителиальных клетках молочной железы крупного рогатого скота. J. Dairy Sci. 103, 6661–6671. дои: 10.3168/jds.2019-17415
дои: 10.3168/jds.2019-17415
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Каука, М., Зикмунд, Т., Тесарова, М., Гилборг, Д., Хелландер, А., Ярос, Дж., и соавт. (2017). Ориентированная динамика клональных клеток обеспечивает точный рост и формирование хрящей позвоночных. Элиф 6:25902. doi: 10.7554/eLife.25902
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кирнан, Дж. А. (2001). Протокол окрашивания Sirius Red для коллагена .Лондон: Университет Западного Онтарио.
Академия Google
Knutsen, G., Drogset, J.O., Engebretsen, L., Grøntvedt, T., Isaksen, V., Ludvigsen, T.C., et al. (2007). Рандомизированное исследование, сравнивающее имплантацию аутологичных хондроцитов с микропереломами. Выводы через пять лет. J. Хирург суставов костей. Являюсь. 89, 2105–2112. doi: 10.2106/JBJS.G.00003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кумар Ю.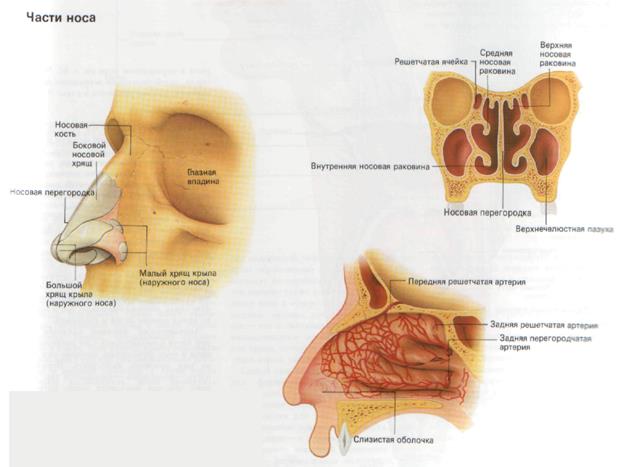 , Бисвас Т., Такер Г., Kanaujiya, J.K., Kumar, S., Shukla, A., et al. (2018). Остеогенез, управляемый передачей сигналов BMP, критически зависит от опосредованного экспрессией Prdx-1 поддержания прегипетрофии хондроцитов. Свободный радикал. биол. Мед. 118, 1–12. doi: 10.1016/j.freeradbiomed.2018.02.016
, Бисвас Т., Такер Г., Kanaujiya, J.K., Kumar, S., Shukla, A., et al. (2018). Остеогенез, управляемый передачей сигналов BMP, критически зависит от опосредованного экспрессией Prdx-1 поддержания прегипетрофии хондроцитов. Свободный радикал. биол. Мед. 118, 1–12. doi: 10.1016/j.freeradbiomed.2018.02.016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лаверна, Л., Браун, У. Э., Вонг, Б. Дж. Ф., Ху, Дж. К., и Афанасиу, К. А. (2019). К тканевой инженерии носовых хрящей. Акта Биоматер. 88, 42–56. doi: 10.1016/j.actbio.2019.02.025
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ли, С.-Ю., Абель, Э.Д., и Лонг, Ф. (2018). Метаболизм глюкозы, индуцируемый передачей сигналов Bmp, необходим для развития скелета мышей. Нац. коммун. 9:4831. doi: 10.1038/s41467-018-07316-5
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лиан, Дж. Б., Макки, М. Д., Тодд, А. М., и Герстенфельд, Л. К. (1993). Индукция костных белков, остеокальцина и остеопонтина, и их ультраструктурная локализация в матриксе с развитием гипертрофии хондроцитов in vitro. J. Cell Biochem. 52, 206–219. doi: 10.1002/jcb.240520212
М., и Герстенфельд, Л. К. (1993). Индукция костных белков, остеокальцина и остеопонтина, и их ультраструктурная локализация в матриксе с развитием гипертрофии хондроцитов in vitro. J. Cell Biochem. 52, 206–219. doi: 10.1002/jcb.240520212
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лю, К., Цзэн, Х., Юань, Ю., Ван, З., Ву, З. и Луо, В. (2020). Остеопонтин ингибирует прогрессирование остеоартрита через сигнальную ось OPN/CD44/PI3K. Гены Дис. 2020:006. doi: 10.1016/j.gendis.2020.06.006
Полнотекстовая перекрестная ссылка | Академия Google
Лю С.-С., Чжоу П. и Чжан Ю. (2016). Аномальная экспрессия ключевых генов и белков в каноническом пути Wnt/β-катенина суставного хряща в крысиной модели остеоартрита, вызванного физической нагрузкой. Мол. Мед. Респ. 13, 1999–2006 гг. doi: 10.3892/mmr.2016.4798
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Луо, П., Гао, Ф. , Ниу, Д., Сунь, X., Сонг, К., Го, К., и другие. (2019). Роль аутофагии в метаболизме хондроцитов и остеоартрозе: всесторонний обзор исследований. Биомед. Рез. Междунар. 2019:5171602. дои: 10.1155/2019/5171602
, Ниу, Д., Сунь, X., Сонг, К., Го, К., и другие. (2019). Роль аутофагии в метаболизме хондроцитов и остеоартрозе: всесторонний обзор исследований. Биомед. Рез. Междунар. 2019:5171602. дои: 10.1155/2019/5171602
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Малик З., Алексиу М., Халлгримссон, Б., Экономидес, А. Н., Людер, Х. У., и Граф, Д. (2018). Костный морфогенетический белок 2 координирует раннюю минерализацию зубов. Дж. Дент. Рез. 97, 835–843. дои: 10.1177/0022034518758044
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Малик З., Рот Д. М., Итон Ф., Теодор Дж. М. и Граф Д. (2020). Мезенхимальный Bmp7 контролирует начало минерализации зубов: новый способ регулирования формы бугорка моляра. Фронт. Физиол. 11:00698.doi: 10.3389/fphys.2020.00698
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мандур, Ю.М.Х., Або Юссеф, С.М., и Мусса, Х. Х. (2019). Изменения полисомнографической и легочной функции у больных с нарушениями сна после септопластики с турбинэктомией. утра. Дж. Отоларингол. 40, 187–190. doi: 10.1016/j.amjoto.2018.12.003
Х. (2019). Изменения полисомнографической и легочной функции у больных с нарушениями сна после септопластики с турбинэктомией. утра. Дж. Отоларингол. 40, 187–190. doi: 10.1016/j.amjoto.2018.12.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мендес-Мальдонадо, К., Вега-Лопес, Г., Кабальеро-Чакон, С., Айбар, М.Дж., и Веласко, И. (2018). Активация Hes1 и Msx1 в эмбриональных стволовых клетках трансгенных мышей увеличивает дифференцировку в производные нервного гребня. Междунар. Дж. Мол. науч. 19:ijms1
25. дои: 10.3390/ijms1
25
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Меррихью, К., Кумар, Б., Эретис, К., Рюгер, Д.С., Кюттнер, К.Е., и Чубинская, С. (2003). Изменения эндогенного остеогенного белка-1 при дегенерации суставного хряща человека. Дж. Ортоп. Рез. 21, 899–907. doi: 10.1016/S0736-0266(03)00055-X
Полнотекстовая перекрестная ссылка | Академия Google
Ми, Х. , Муругануджан, А., Эберт, Д., Хуанг, X., и Томас, П. Д. (2019). PANTHER версии 14: больше геномов, новый PANTHER GO-slim и улучшения в инструментах анализа обогащения. Рез. нуклеиновых кислот. 47, Д419–Д426. doi: 10.1093/nar/gky1038
, Муругануджан, А., Эберт, Д., Хуанг, X., и Томас, П. Д. (2019). PANTHER версии 14: больше геномов, новый PANTHER GO-slim и улучшения в инструментах анализа обогащения. Рез. нуклеиновых кислот. 47, Д419–Д426. doi: 10.1093/nar/gky1038
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Могхаддам, М., Гарсия, Г.Дж. М., Франк-Ито, Д. О., Кимбелл, Дж. С., и Ри, Дж. С. (2020). Виртуальная септопластика: метод прогнозирования хирургических результатов у пациентов с обструкцией носовых дыхательных путей. Междунар. Дж. АВТОМОБИЛИ 15, 725–735. doi: 10.1007/s11548-020-02124-z
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Морита К., Миямото Т., Фудзита Н., Кубота Ю., Ито К., Такубо К. и др. (2007). Активные формы кислорода вызывают гипертрофию хондроцитов при эндохондральном окостенении. Дж.Эксп. Мед. 204, 1613–1623. doi: 10.1084/jem.20062525
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мрачек Т. , Драхота З. и Хоуштек Дж. (2013). Функция и роль митохондриальной глицерол-3-фосфатдегидрогеназы в тканях млекопитающих. Биохим. Биофиз. Акта Биоэнергет. 1827, 401–410. doi: 10.1016/j.bbabio.2012.11.014
, Драхота З. и Хоуштек Дж. (2013). Функция и роль митохондриальной глицерол-3-фосфатдегидрогеназы в тканях млекопитающих. Биохим. Биофиз. Акта Биоэнергет. 1827, 401–410. doi: 10.1016/j.bbabio.2012.11.014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мумме, М., Барберо, А., Miot, S., Wixmerten, A., Feliciano, S., Wolf, F., et al. (2016). Сконструированная на основе носовых хондроцитов аутологичная хрящевая ткань для восстановления дефектов суставного хряща: первое обсервационное испытание на людях. Ланцет 388, 1985–1994 гг. дои: 10.1016/S0140-6736(16)31658-0
Полнотекстовая перекрестная ссылка | Академия Google
Музумдар, М. Д., Тасич, Б., Миямичи, К., Ли, Л., и Луо, Л. (2007). Глобальная репортерная мышь Cre с двойной флуоресценцией. Бытие 45, 593–605.doi: 10.1002/dvg.20335
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нисимура К., Мурозуми К., Ширахата А. , Пак М. Х., Кашиваги К. и Игараши К. (2005). Независимые роли eIF5A и полиаминов в пролиферации клеток. Биохим. J. 385, 779–785. дои: 10.1042/BJ20041477
, Пак М. Х., Кашиваги К. и Игараши К. (2005). Независимые роли eIF5A и полиаминов в пролиферации клеток. Биохим. J. 385, 779–785. дои: 10.1042/BJ20041477
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Санчес К., Бэй-Дженсен А.-К., Пап Т., Двир-Гинзберг М., Квасничка Х., Барретт-Джолли Р., и другие. (2017). Секретом хондроцитов: источник новых идей и поисковых биомаркеров остеоартрита. Остеоартроз. Картил. 25, 1199–1209. doi: 10.1016/j.joca.2017.02.797
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Скьявинато, А., Беккер, А.-К. А., Дзанетти М., Коралло Д., Миланетто М., Биззотто Д. и др. (2012). EMILIN-3, своеобразный член семейства белков, локализованных на поверхности микрофибрилл эластина (EMILIN), имеет четкий паттерн экспрессии, образует олигомерные сборки и служит антагонистом трансформирующего фактора роста β (TGF-β). Дж. Биол. хим. 287, 11498–11515. doi: 10.1074/jbc.M111. 303578
303578
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шульц Т.Дж. и Ценг Ю.-Х. (2009). Новая роль костных морфогенетических белков в адипогенезе и энергетическом обмене. Фактор роста цитокинов Ред. 20, 523–531. doi: 10.1016/j.cytogfr.2009.10.019
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сеол, Д., Маккейб, Д. Дж., Чоу, Х., Чжэн, Х., Yu, Y., Jang, K., et al. (2012). Хондрогенные клетки-предшественники реагируют на повреждение хряща. Ревматоидный артрит. 64, 3626–3637. дои: 10.1002/арт.34613
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Серифоглу И., Оз И.И., Дамар М., Буюкуйсал М.С., Тосун А. и Токгоз О. (2017). Взаимосвязь между степенью и направлением искривления носовой перегородки и морфологией костей носа. Head Face Med. 13:3. doi: 10.1186/s13005-017-0136-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тагилу, Х.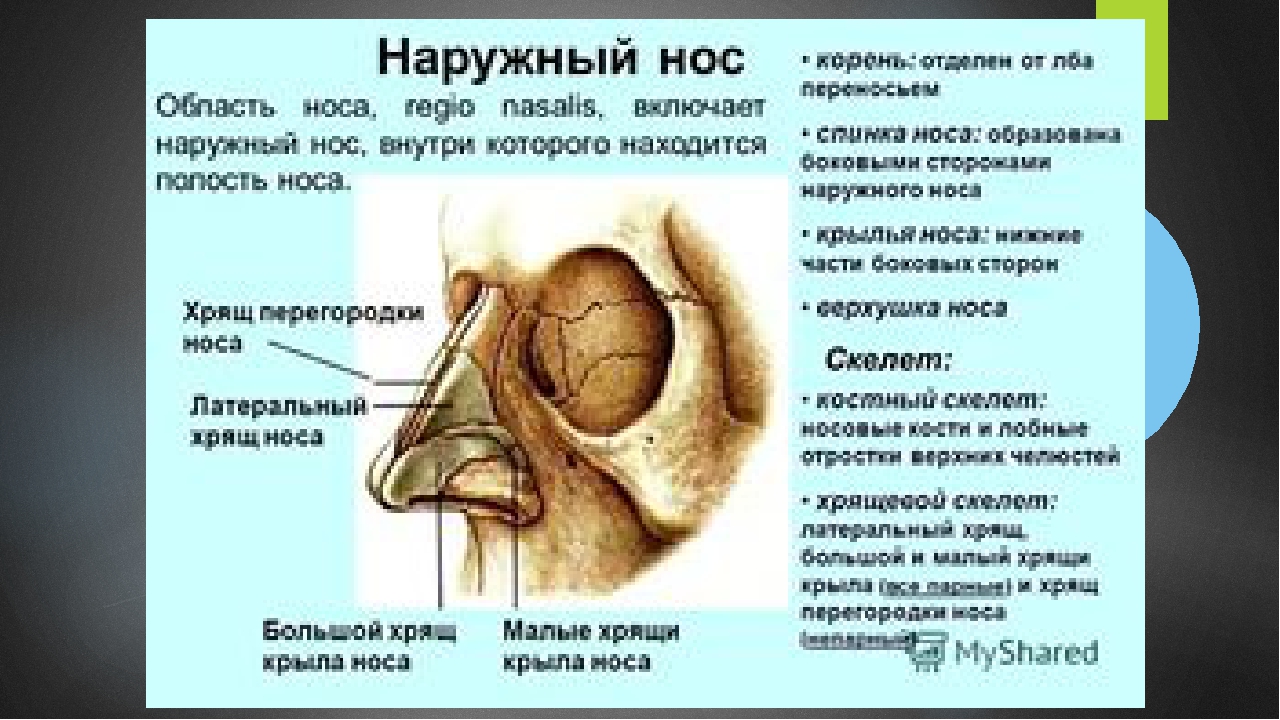 и Халими, З. (2019). Частота различных видов искривления носовой перегородки и их влияние на увеличение толщины слизистой оболочки верхнечелюстной пазухи. Дж. Дент. Рез. Вмятина. клин. Вмятина. Проспекты 13, 208–214. doi: 10.15171/joddd.2019.032
и Халими, З. (2019). Частота различных видов искривления носовой перегородки и их влияние на увеличение толщины слизистой оболочки верхнечелюстной пазухи. Дж. Дент. Рез. Вмятина. клин. Вмятина. Проспекты 13, 208–214. doi: 10.15171/joddd.2019.032
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Четина Е.В., Маркова Г.А. (2018). Регуляция энергетического обмена в пластинке роста и остеоартритных хондроцитах. Ревматол. Междунар. 38, 1963–1974 гг.doi: 10.1007/s00296-018-4103-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тейшейра Дж., Сертал В., Чанг Э. Т. и Камачо М. (2016). Отклонения носовой перегородки: систематический обзор систем классификации. Пласт. Surg. Междунар. 2016:7089123. дои: 10.1155/2016/7089123
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Цолис, К.С., Бей, Э.С., Папатанасиу, И., Костопулу, Ф., Гкреци, В., Каланцаки, К.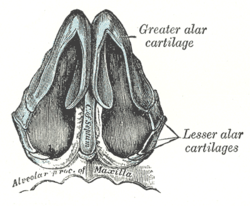 , и соавт.(2015). Сравнительный протеомный анализ гипертрофированных хондроцитов при остеоартрозе. клин. Протеомика 12:12. doi: 10.1186/s12014-015-9085-6
, и соавт.(2015). Сравнительный протеомный анализ гипертрофированных хондроцитов при остеоартрозе. клин. Протеомика 12:12. doi: 10.1186/s12014-015-9085-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
ван дер Краан, П.М., и ван ден Берг, В.Б. (2012). Гипертрофия хондроцитов и остеоартроз: роль в инициации и прогрессировании дегенерации хряща? Остеоартрит. Картил. 20, 223–232. doi: 10.1016/j.joca.2011.12.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
ван Лисхаут, Т.Л. и Любичич В. (2019). Появление белков аргининметилтрансфераз в скелетных мышцах и нарушениях обмена веществ. утра. Дж. Физиол. Эндокринол. Метабол. 317, Е1070–Е1080. doi: 10.1152/ajpendo.00251.2019
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вора, С.Р., Камчи, Э.Д., и Кокс, Т.С. (2015). Постнатальный онтогенез основания черепа и черепно-лицевого скелета у самцов мышей C57BL/6J: эталонный стандарт для количественного анализа. Фронт.Физиол. 6:417. doi: 10.3389/fphys.2015.00417
Фронт.Физиол. 6:417. doi: 10.3389/fphys.2015.00417
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Wang, J., Dou, X., Liu, D., Song, P., Qian, X., Wang, S., et al. (2016). Оценка влияния искривления носовой перегородки на строение полости носа. евро. Арка Оториноларингол. 273, 1477–1480. doi: 10.1007/s00405-015-3770-y
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уэлтхолл, Р. Дж., и Херринг, С.В. (2006). Эндохондральная оссификация носовой перегородки мыши. Анат. Рек. Дисков. Мол. Клетка Эвол. биол. 288, 1163–1172. doi: 10.1002/ar.a.20385
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вонг, М.К., Шоуки, С.А., Арьясомайюла, А., Грин, М.А., Юарт, Т., Селваганапати, П.Р., и соавт. (2018). Поверхность внеклеточного матрикса регулирует самосборку трехмерных сфероидов плацентарного трофобласта. PLoS One 13:0199632. дои: 10.1371/journal.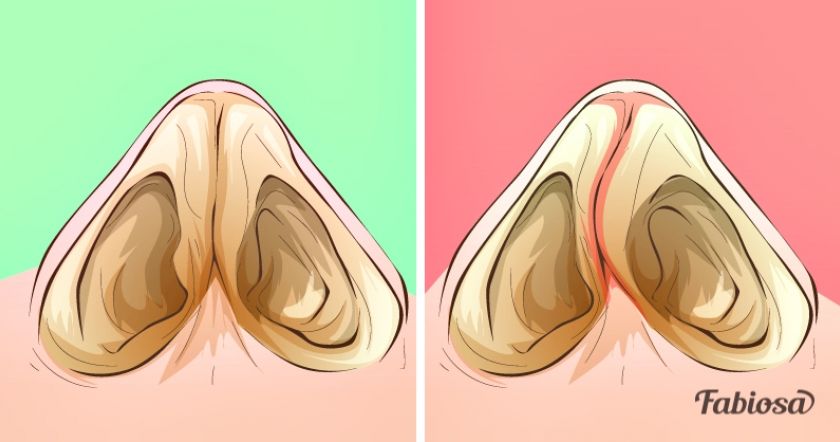 pone.0199632
pone.0199632
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сюй Д., Джин Т., Чжу Х., Чен Х., Офенгейм Д., Цзоу С. и др. (2018). TBK1 подавляет апоптоз и воспаление, управляемые RIPK1, во время развития и старения. Ячейка 174, 1477.e–1491.e. doi: 10.1016/j.cell.2018.07.041
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ян, М., Араи, А., Удагава, Н., Хирага, Т., Лиджуан, З., Ито, С., и др. (2017).Остеогенный фактор Runx2 маркирует подмножество клеток, положительных по рецептору лептина, которые находятся на вершине иерархии стромальных клеток костного мозга. науч. Респ. 7:4928. doi: 10.1038/s41598-017-05401-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Йошинари, К., Сато, Т., Окино, Н., Сугатани, Дж., и Мива, М. (2004). Экспрессия и индукция цитохромов р450 в белой жировой ткани крыс. Дж. Фармакол. Эксп. тер. 311, 147–154. doi: 10.1124/jpet. 104.067066
104.067066
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ю, Дж.С.Л. и Цуй В. (2016). Пролиферация, выживание и метаболизм: роль передачи сигналов PI3K/AKT/mTOR в определении плюрипотентности и судьбы клеток. Разработка 143, 3050–3060. doi: 10.1242/dev.137075
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Yu, X., Kawakami, H., Tahara, N., Olmer, M., Hayashi, S., Akiyama, R., et al. (2017). Экспрессия Noggin и Gremlin1 и ее значение для тонкой настройки активности BMP в хрящевых тканях мыши. Дж.Ортоп. Рез. 35, 1671–1682. doi: 10.1002/jor.23463
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зеленски, Н. А., Ледди, Х. А., Санчес-Адамс, Дж., Чжан, Дж., Бональдо, П., Лидтке, В., и соавт. (2015). Коллаген VI регулирует свойства перицеллюлярного матрикса, набухание хондроцитов и механотрансдукцию в суставном хряще. Артрит Ревматолог. 67, 1286–1294. дои: 10.1002/арт.39034
дои: 10.1002/арт.39034
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжэн, М.-H., Willers, C., Kirilak, L., Yates, P., Xu, J., Wood, D., et al. (2007). Индуцированная матриксом имплантация аутологичных хондроцитов (MACI): биологическая и гистологическая оценка. Ткань Eng. 13, 737–746. doi: 10.1089/ten.2006.0246
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжун, Л., Хуанг, X., Карпериен, М., и Пост, Дж. Н. (2016a). Корреляция между экспрессией генов и прогрессированием остеоартрита у человека. Междунар. Дж. Мол. науч. 17:ijms17071126.дои: 10.3390/ijms17071126
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Zhong, L., Huang, X., Rodrigues, E.D., Leijten, J.C.H., Verrips, T., El Khattabi, M., et al. (2016б). Эндогенные DKK1 и FRZB регулируют хондрогенез и гипертрофию в трехмерных культурах хондроцитов человека и мезенхимальных стволовых клеток человека. Разработка стволовых клеток. 25, 1808–1817 гг. doi: 10.1089/scd.2016.0222
Разработка стволовых клеток. 25, 1808–1817 гг. doi: 10.1089/scd.2016.0222
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Носовой скелет – кости – хрящи – переломы
Скелет носа представляет собой комбинацию костей и хрящей, формирующих то, что мы можем видеть как внешний нос, и внутреннюю носовую перегородку, которая разделяет две носовые полости головы.
Здесь мы обсудим анатомию носового скелета и составляющих его костей.
Анатомическая структура
Скелет носа образован тремя типами тканей; кости, хряща и фиброзно-жировой ткани. При взгляде на каркас носа полезно разделить конструкции на две части; наружный носовой скелет и внутренняя носовая перегородка .
Внешний скелет носа
Рисунок 1. Внешний вид скелета носа, вид сбоку Внешний скелет расширяет носовые полости на переднюю часть лица (см.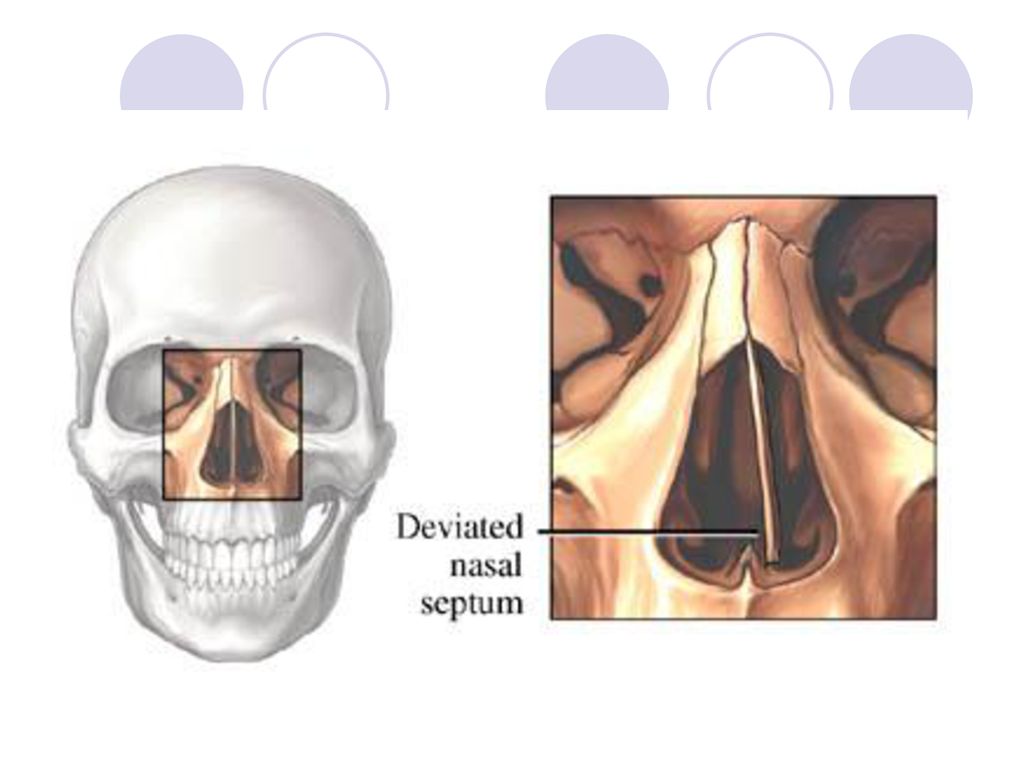 рис. 1).Он частично образован носовой и верхнечелюстной костями, расположенными выше. Нижняя часть носа состоит из гиалиновых хрящей; боковые, большие крылья, малые крылья и хрящевая перегородка . Боковые и основные крыловые хрящи являются самыми большими и вносят наибольший вклад в форму носа. Малые крыловые хрящи различаются по количеству, обычно их 3 или 4 с каждой стороны.
рис. 1).Он частично образован носовой и верхнечелюстной костями, расположенными выше. Нижняя часть носа состоит из гиалиновых хрящей; боковые, большие крылья, малые крылья и хрящевая перегородка . Боковые и основные крыловые хрящи являются самыми большими и вносят наибольший вклад в форму носа. Малые крыловые хрящи различаются по количеству, обычно их 3 или 4 с каждой стороны.
Внутренняя носовая перегородка
Внутренняя носовая перегородка разделяет полость носа на две ноздри.Кости, входящие в состав носовой перегородки, можно разделить на:
- Парные кости : Носовые, верхнечелюстные и небные кости
- Непарные кости : Решетчатая и сошниковая кости.
В дополнение к костям носа, перегородки и большие крыловидные хрящи также составляют часть носовой перегородки.
Решетчатая кость участвует в центральной части носовой перегородки.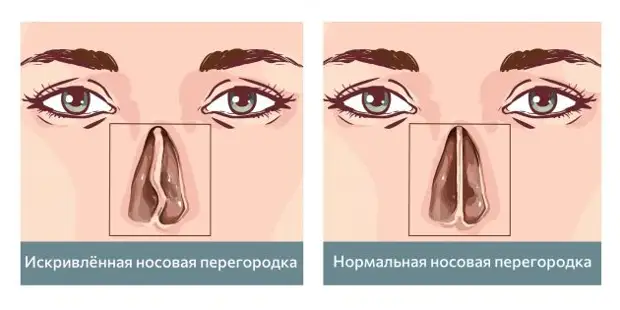 Это одна из самых сложных костей в человеческом теле, и ее строение выходит за рамки этой статьи, однако дополнительную информацию можно найти здесь.Передняя и задняя части образованы септальным хрящом и сошником костью соответственно.
Это одна из самых сложных костей в человеческом теле, и ее строение выходит за рамки этой статьи, однако дополнительную информацию можно найти здесь.Передняя и задняя части образованы септальным хрящом и сошником костью соответственно.
Пол полости носа образован твердым небом, отделяющим его от полости рта. Твердое небо состоит из небной кости сзади и небного отростка верхней челюсти спереди. Решетчатая пластинка решетчатой кости образует крышу полости носа.
Рисунок 2. Боковой вид носовой перегородки сбоку.[/подпись][старт-клинический]
Клиническая значимость: перелом носа
Из-за выступа наружного скелета носа часто встречаются переломы носа — наиболее распространен перелом лица. Переломы обычно возникают в результате тупой травмы носа. Обычным последствием переломов носа является необратимая деформация из-за разрушения костей и хрящей.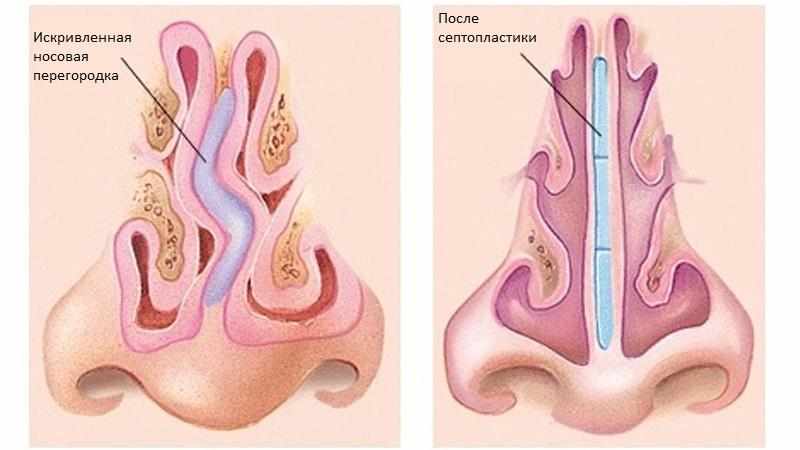
[конечный клинический]
Гистологическая и молекулярная характеристика растущей носовой перегородки у мышей
Носовая перегородка представляет собой хрящевую структуру, которая служит водителем ритма для развития средней зоны лица.Перегородка представляет собой гиалиновый хрящ, окруженный надхрящницей и эпителием. Спереди она остается хрящевой, но сзади подвергается эндохондральному окостенению, образуя перпендикулярную пластинку решетчатой кости. Понимание дифференцировки гиалинового хряща связано преимущественно с исследованиями хряща пластинки роста. В настоящее время неясно, совпадают ли морфологические и молекулярные свойства дифференцирующейся носовой перегородки с тем, что известно из пластинки роста.В этом исследовании мы описываем рост, молекулярные и клеточные характеристики носовой перегородки со ссылкой на дифференцировку гиалинового хряща. Носовая перегородка растет асинхронно по всей длине, причем фазы быстрого роста прерываются более застойным ростом. Рост, по-видимому, обусловлен преимущественно приобретением гипертрофии хондроцитов. Точно так же клеточная дифференцировка асинхронна, и дифференцировка, наблюдаемая в передней части, предшествует задней дифференцировке. В целом носовая перегородка структурно и молекулярно неоднородна.Ранняя и обширная гипертрофия хондроцитов, но не наблюдается окостенения в передней части перегородки. Начало гипертрофической дифференцировки хондроцитов совпало с отложением коллагеновых волокон вдоль надхрящницы. Экспрессия Sox9, Col2, Col10, Mmp13, Sp7 и Runx2 была гетерогенной и не всегда соответствовала ожидаемому паттерну, установленному из дифференцировки хондроцитов в ростовой пластинке. Присутствие гипертрофированных хондроцитов, экспрессирующих белки, связанные с костью, на ранних стадиях в областях, где носовая перегородка не окостеневает, демонстрирует несоответствие с современным пониманием дифференцировки гиалинового хряща.Runx2, Collagen II, Collagen X и Sp7, обычно используемые для обозначения различных стадий созревания хондроцитов и раннего формирования кости, демонстрируют более широкую экспрессию, чем ожидалось, и не соответствуют ожидаемым клеточным характеристикам.
Рост, по-видимому, обусловлен преимущественно приобретением гипертрофии хондроцитов. Точно так же клеточная дифференцировка асинхронна, и дифференцировка, наблюдаемая в передней части, предшествует задней дифференцировке. В целом носовая перегородка структурно и молекулярно неоднородна.Ранняя и обширная гипертрофия хондроцитов, но не наблюдается окостенения в передней части перегородки. Начало гипертрофической дифференцировки хондроцитов совпало с отложением коллагеновых волокон вдоль надхрящницы. Экспрессия Sox9, Col2, Col10, Mmp13, Sp7 и Runx2 была гетерогенной и не всегда соответствовала ожидаемому паттерну, установленному из дифференцировки хондроцитов в ростовой пластинке. Присутствие гипертрофированных хондроцитов, экспрессирующих белки, связанные с костью, на ранних стадиях в областях, где носовая перегородка не окостеневает, демонстрирует несоответствие с современным пониманием дифференцировки гиалинового хряща.Runx2, Collagen II, Collagen X и Sp7, обычно используемые для обозначения различных стадий созревания хондроцитов и раннего формирования кости, демонстрируют более широкую экспрессию, чем ожидалось, и не соответствуют ожидаемым клеточным характеристикам.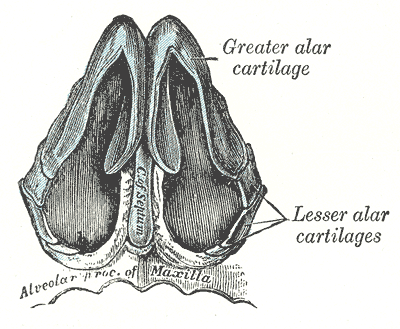 Таким образом, гиалиновый хрящ носовой перегородки весьма отличается от гиалинового хряща пластинки роста, и следует соблюдать осторожность, прежде чем приписывать свойства хряща менее четко определенным хрящевым структурам с использованием этих обычно используемых маркеров. Помимо структурного описания носового хряща, это исследование также предоставляет важную информацию для инженерии хрящевой ткани при использовании хряща носовой перегородки для регенерации тканей.
Таким образом, гиалиновый хрящ носовой перегородки весьма отличается от гиалинового хряща пластинки роста, и следует соблюдать осторожность, прежде чем приписывать свойства хряща менее четко определенным хрящевым структурам с использованием этих обычно используемых маркеров. Помимо структурного описания носового хряща, это исследование также предоставляет важную информацию для инженерии хрящевой ткани при использовании хряща носовой перегородки для регенерации тканей.
Ключевые слова: маркеры дифференцировки хондроцитов; внеклеточный матрикс; развитие гиалинового хряща; мышь; носовая перегородка.
Клинический анализ хряща носовой перегородки с помощью компьютерной томографии
Arch Craniofac Surg. 2016 сен; 17(3): 140–145.
, , , , иСо Мин Хван
Корейский центр эстетической, пластической и реконструктивной хирургии, Busanh Hospital, Good Moonh.
Он Лим
Центр эстетической, пластической и реконструктивной хирургии, больница Гуд Мунхва, Пусан, Корея.
Мин Кю Хван
Центр эстетической, пластической и реконструктивной хирургии, больница Гуд Мунхва, Пусан, Корея.
Мин Вук Ким
Центр эстетической, пластической и реконструктивной хирургии, больница Гуд Мунхва, Пусан, Корея.
Jong Seo Lee
Центр эстетической, пластической и реконструктивной хирургии, больница Good Moonhwa, Пусан, Корея.
Центр эстетической, пластической и реконструктивной хирургии, больница Гуд Мунхва, Пусан, Корея.
Автор, ответственный за переписку. Переписка: Со Мин Хван. Центр эстетической, пластической и реконструктивной хирургии, больница Гуд Мунхва, 119 Беомил-ро, Донг-гу, Пусан 48735, Корея. ten.liamnah@gnawhnimosПоступила в редакцию 25 марта 2016 г.; Пересмотрено 12 сентября 2016 г.; Принято 12 сентября 2016 г.
Copyright © Корейская ассоциация расщелин неба и черепно-лицевой области, 2016 г.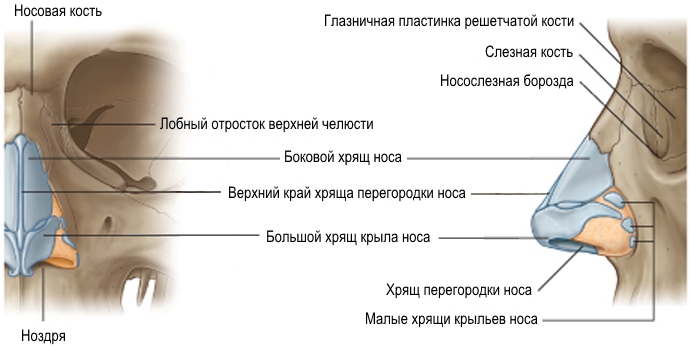 org/licenses/by-nc/3.0/), что разрешает неограниченное некоммерческое использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы. Эта статья цитировалась в других статьях PMC.
org/licenses/by-nc/3.0/), что разрешает неограниченное некоммерческое использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы. Эта статья цитировалась в других статьях PMC.Abstract
История вопроса
Хрящ перегородки носа часто используется в качестве донорского трансплантата при операциях по ринопластике, но его размер может широко варьироваться в зависимости от группы пациентов. Таким образом, предоперационная оценка хрящевой области важна как для консультирования пациентов, так и для планирования операции.Мы стремимся оценить площадь септального хряща с помощью компьютерной томографии (КТ) лица.
Методы
Исследование проводилось с использованием КТ-изображений лица, полученных у 200 пациентов в период с января 2012 г. по июль 2015 г. На срединно-сагиттальном изображении граница хрящевой перегородки была отграничена от мягких тканей с использованием средней разницы в интенсивности сигнала (или яркость). Площадь в пределах этой границы была рассчитана. Затем вычисленная площадь септального хряща сравнивалась для разных возрастных групп и полов.
Площадь в пределах этой границы была рассчитана. Затем вычисленная площадь септального хряща сравнивалась для разных возрастных групп и полов.
Результаты
В целом средняя площадь хряща носовой перегородки составила 8,18 см 2 с максимальной площадью 12,42 см 2 и минимальной площадью 4,89 см 2 . Было измерено, что площади хрящей больше у мужчин, чем у женщин ( p <0,05). Площадь уменьшалась с возрастом ( p <0,05).
Заключение
Измерение размера септального хряща с использованием разницы яркости является более точным и надежным, чем описанные ранее методы.Этот метод может быть использован как стандарт для профилактики послеоперационных осложнений. Носовой хрящ, компьютерная томография, измерение Клинически септальный хрящ важен, поскольку он используется в качестве донорского хряща для многих операций по ринопластике [1,2].Объем носовой перегородки, занятой хрящевым компонентом, различен у разных пациентов, что приводит к большим вариациям формы и размера хряща носовой перегородки.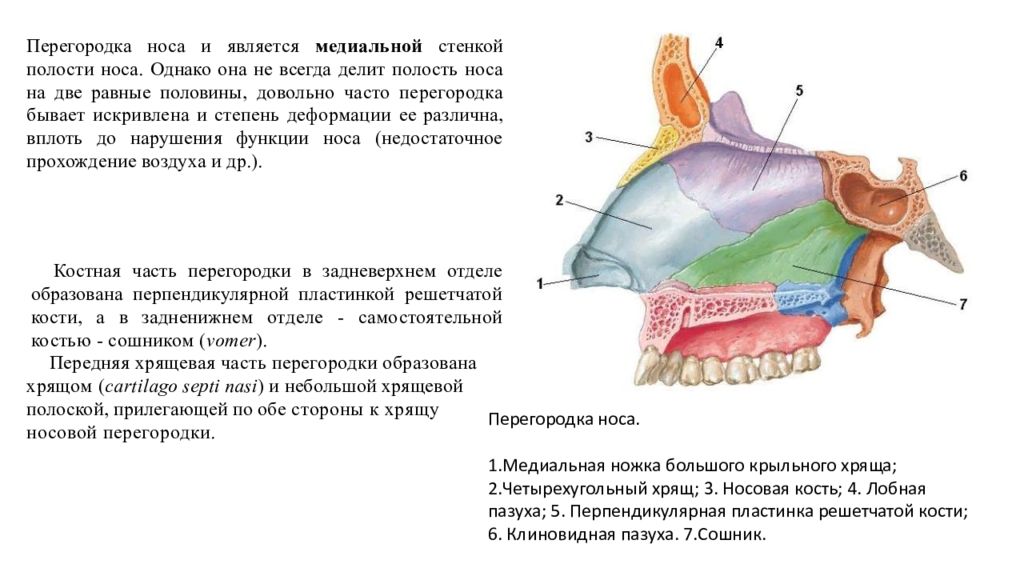 Следовательно, важно оценить количество септального хряща, доступного для сбора. Тем не менее, не существует стандартного метода предоперационной оценки сбора септального хряща.
Следовательно, важно оценить количество септального хряща, доступного для сбора. Тем не менее, не существует стандартного метода предоперационной оценки сбора септального хряща.
В предыдущих исследованиях размеры хрящей носовой перегородки изучались на трупах с помощью магнитно-резонансной томографии (МРТ) и компьютерной томографии (КТ) [3,4,5,6].Хотя трупные измерения точны, данные не имеют отношения к клиническим условиям [3,4]. Измерение МРТ полезно для изучения различий между хрящом носовой перегородки и мягкими тканями. Однако высокая стоимость МРТ не позволяет широко использовать ее для эстетической ринопластики, поэтому данные МРТ очень редко доступны в соответствующих клинических условиях [5]. С другой стороны, КТ является наиболее часто используемым методом для выявления перелома и искривления носовой перегородки при травме лица и является относительно недорогим по сравнению с МРТ методом при самой высокой клинической доступности [1].Кроме того, яркость между тканями можно сравнивать только с помощью простой работы программы, что позволяет более точно оценить размеры септального хряща.
В этом исследовании мы объективно измерили разницу в яркости между подкожной тканью и хрящом с помощью программного решения. На основании данных яркости мы смогли очертить границы септального хряща. Эта информация может быть важна при предоперационном планировании забора септального хряща и может использоваться в качестве справочных данных для предотвращения осложнений, таких как послеоперационные деформации носа.
МЕТОДЫ
Это исследование было одобрено Институциональным наблюдательным советом Университета Индже (№ 2015172-2) и соответствовало Кодексу этики научных исследований.
Это ретроспективное исследование было проведено для пациентов, которые посещали нашу клинику и проходили компьютерную томографию в течение 3,5 лет с января 2012 г. по июль 2015 г. и группа подростков. Также были исключены пациенты с тяжелым искривлением перегородки или наличием перелома перегородки.
КТ-изображения получены при 120 кВ и 180 мА. Для измерения площади сначала устанавливалась точка, где были разделены оба полушария головного мозга. Впоследствии сагиттальные изображения были извлечены с использованием программного обеспечения PiViewSTAR (программное обеспечение INFINITT, Сеул, Корея). Границы хряща были отмечены на извлеченном изображении перед измерением площади.
Впоследствии сагиттальные изображения были извлечены с использованием программного обеспечения PiViewSTAR (программное обеспечение INFINITT, Сеул, Корея). Границы хряща были отмечены на извлеченном изображении перед измерением площади.
Мы использовали разницу яркости между тканями при маркировке границы, чтобы обеспечить объективность в различении ткани септального хряща от подкожной ткани носа.Этот метод в основном используется для неразрушающих испытаний и измеряет разницу в яркости, создаваемую между различными тканями, которую можно подтвердить с помощью существующей программы (). На каждом КТ-снимке выбирали по 3 точки, исключая участки, контактирующие с костями. По точкам измеряли разность яркостей, после чего отмечали области, демонстрирующие резкие различия (). С ними в качестве отправных точек была отмечена граница области.
(A) Установка средней линии и измерение площади септального хряща.Срединная сагиттальная плоскость использовалась в качестве плоскости отсчета.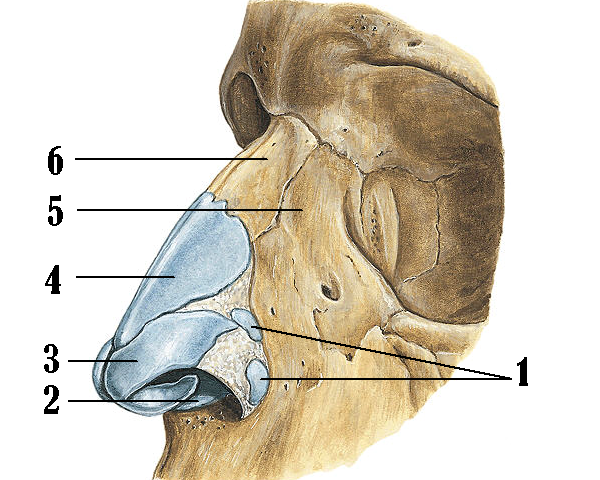 (B) Среднесагиттальное изображение компьютерной томографии. Выявление разницы яркости между хрящевой тканью и подкожной клетчаткой. Гистограмму яркости можно использовать для различения тканей.
(B) Среднесагиттальное изображение компьютерной томографии. Выявление разницы яркости между хрящевой тканью и подкожной клетчаткой. Гистограмму яркости можно использовать для различения тканей.
(A) Мы установили одну точку на изображении между самым передним кончиком носа и нижним краем носовой кости. По точкам измеряли разницу яркости между подкожной клетчаткой и септальным хрящом.И мы отметили край септального хряща. (Б) На компьютерной томографии были выбраны 3 точки, исключая участки, контактирующие с костями. А — срединная точка между точкой В и нижним краем носовой кости; B, самый передний конец септального хряща; C, самая нижняя точка линии шва между септальным хрящом и перпендикулярной пластинкой решетчатой кости.
Затем на извлеченном изображении была отмечена граница септального хряща с использованием программного обеспечения GoodNotes (Time Base Technology, Quarry Bay, Hongkong), а затем граница хряща, прилегающая к перпендикулярной пластинке решетчатой кости и сошника. Впоследствии площадь была измерена с использованием программного обеспечения Paint.net (dotPDN, Олимпия, Вашингтон, США) (). Мы измеряли площадь и яркость для всех изображений в каждом исследовании. Все измерения проводились дважды и усреднялись.
Впоследствии площадь была измерена с использованием программного обеспечения Paint.net (dotPDN, Олимпия, Вашингтон, США) (). Мы измеряли площадь и яркость для всех изображений в каждом исследовании. Все измерения проводились дважды и усреднялись.
Граница септального хряща (красная линия).
Статистический анализ был выполнен с использованием статистического программного обеспечения MedCalc ver. 16.4.3 (MedCalc Software bvba, Остенде, Бельгия; https://www.medcalc.org; 2016). Для определения различий были проведены t-критерий независимых выборок и критерий дисперсионного анализа.Различия считались статистически значимыми, когда значение p было меньше 0,05.
РЕЗУЛЬТАТЫ
Обзор выявил 200 пациентов, отвечающих критериям включения (136 мужчин и 64 женщины). Средний возраст составил 37,1 года. Повреждение лица было наиболее частым показанием к КТ лица. Другие показания включали заложенность носа и предоперационную оценку бимаксиллярной хирургии.
Средняя яркость подкожной клетчатки составила 3,83 в диапазоне от -45 до 44 со стандартным отклонением 12.12. Средняя яркость септального хряща составила 54,56, в диапазоне от 4 до 75, со стандартным отклонением 10,87. Следовательно, средняя разница между двумя тканями составила 50,73 в диапазоне от 8 до 87 со стандартным отклонением 7,34.
Средняя площадь септального хряща составила 8,18 см 2 в диапазоне от 4,89 до 12,42 см 2 со стандартным отклонением 1,38. У мужчин средняя площадь септального хряща составляла 8,57 см 2 , тогда как у женщин она составляла 7,36 см 2 , и эта разница была значимой ( p <0.05) (). Что касается возраста, то средняя площадь составила 9,89 см 2 для группы от 18 до 20 лет, 8,68 см 2 для группы от 21 до 30 лет, 8,23 см 2 для группы группа от 31 до 40 лет, 7,79 см 2 для группы от 41 до 50 лет и 7,02 см 2 для группы от 51 до 78 лет. Это уменьшение площади перегородки с увеличением возраста было статистически значимым ( p <0,05) ().
Таблица 1
Площадь септального хряща в зависимости от пола
Таблица 2
Площадь септального хряща в зависимости от возраста косметическая хирургия.Планировалась аугментационная ринопластика с пластикой кончика носа и септальным удлиняющим трансплантатом. Поскольку септальный хрящ планировалось использовать в качестве донорского трансплантата, была проведена компьютерная томография лица для оценки количества хряща, доступного для септального трансплантата (). Используя методы, описанные выше, площадь хряща носовой перегородки была измерена и составила 7,45 см 2 . Чтобы сохранить L-образную стойку, хрящ носовой перегородки был разработан на основе КТ-изображения. Измерение площади без учета L-образной стойки оценивается как 4.78 см 2 . Это считалось достаточным количеством хряща. Затем оператор прочитал расположение хряща, операция прошла без каких-либо затруднений.
Компьютерная томография из представленного случая. Мы смогли измерить собираемое количество хряща носовой перегородки. A+B, тотальный септальный хрящ; B, собираемый хрящ носовой перегородки. Двойные стрелки обозначают смещение в 1 см как ширину L-образной стойки.
ОБСУЖДЕНИЕ
В контексте операций по ринопластике хрящ носовой перегородки часто используется в качестве структурного каркаса в том же операционном поле, что и сама операция [7,8,9].Среди вариантов хрящевого трансплантата для ринопластики реберный хрящ является наиболее удаленным от операционного поля и оставляет рубец после операции. Ушной хрящ имеет низкую жесткость по сравнению с септальным хрящом. Следовательно, септальный хрящ предпочтительнее, когда хрящ должен быть собран для структурного каркаса [9,10].
В дополнение к прямой трансплантации хряща также предпочтителен непрямой метод, при котором уже собранные хондроциты вводят в полигликолевый каркас и культивируют в нем с последующей пересадкой в зону реконструкции.Однако хрящ в основном состоит из коллагена и протеогликанов, а количество хондроцитов у взрослых невелико [7]. Таким образом, хондроциты предпочтительно собирать из переднего и среднего отделов хряща носовой перегородки, где хондроциты наиболее многочисленны и обладают наибольшей жизнеспособностью [11].
Перегородочный хрящ является важной структурой не только для прямой трансплантации, но и для непрямой трансплантации с использованием тканевой инженерии в реконструктивной хирургии.Следовательно, предоперационная оценка считается необходимой при операциях, требующих трансплантации септального хряща. Согласно предыдущим исследованиям, площадь хряща носовой перегородки у трупов составляла 8,4 см 2 у мужчин и 7,9 см 2 у женщин [3], 7,5 см 2 на КТ [7] и 8,9 см 2 у женщин. см 2 на МРТ [5]. Как правило, хрящ носовой перегородки быстро увеличивается в размерах от рождения до подросткового возраста. После подросткового периода часть хряща постепенно окостеневает в процессе старения, и, таким образом, площадь септального хряща уменьшается по отношению ко всей носовой перегородке [1].До сих пор исследования размеров септального хряща у мужчин и женщин дали противоречивые результаты: некоторые из них предполагают значительную разницу, а другие нет [5].
Недавние разработки в области пластики кончика носа с использованием хрящевого трансплантата повысили интерес к септальному хрящу. Однако было трудно оценить анатомические особенности септального хряща до фактической операции. В прошлом КТ имела низкую доступность и не могла разрешить хрящевые области из-за низкого разрешения и проблемы с дозой облучения, что ограничивало использование КТ для операций ринопластики.Следовательно, МРТ была основным инструментом для этой цели [12]. Однако недавний прогресс в технологии КТ привел к дифференциации подкожно-жирового слоя в области носа, и стало легче подтвердить границы септального хряща. В то же время значительно улучшилась клиническая доступность КТ [7]. Кроме того, КТ лица стала обычным методом предоперационного обследования при переломе носа, при котором вправление перелома часто сопровождается косметической операцией.
В исследовании, измерявшем площадь септального хряща с помощью КТ лица, Kim et al.[6] отметили и соединили 7 специфических точек на границе септального хряща и оценили площадь хряща в 7,5 см 2 . В последующем исследовании, проведенном той же группой, площадь септального хряща была более точно измерена путем маркировки самой границы септального хряща. Было обнаружено, что средняя площадь составляет 8,3 см 2 с измеренными площадями в диапазоне от минимума 4,4 см 2 до максимума 14,1 см 2 , что свидетельствует о больших различиях в площади септального хряща в выборке. [1].Поэтому мы использовали разницу яркости между тканями для более точного измерения площади хрящевой перегородки.
Гистологический состав септального хряща составляет 77,7% воды, 7,7% коллагена и 2,9% сульфатированных гликозаминогликанов [7]. Хрящ должен показывать разницу в яркости по сравнению с подкожными тканями из-за различия в этом гистологическом составе, на основе которого мы разработали метод оценки септального хряща.
Мы обнаружили, что средняя яркость подкожной клетчатки равнялась 3.83 с минимальным значением -45 и максимальным значением 44, что свидетельствует о значительных различиях между людьми. Средняя яркость хряща составила 50,73 с минимумом -4 и максимумом 75, что также свидетельствует об индивидуальных различиях. Несмотря на перекрытие интенсивностей между двумя тканями, средняя разница в яркости между подкожной тканью и хрящом у человека составляла 50,73 с минимумом 8 и максимумом 87, что было достаточно, чтобы различить две ткани без обмана.
Используя эти данные, площадь септального хряща сравнивали в зависимости от возраста и пола. Результат показал, что хрящ перегородки носа был больше у мужчин, чем у женщин, и что он начал уменьшаться в размерах после второго десятилетия жизни. Пост-подростковое уменьшение размера септального хряща связано с процессом окостенения, во время которого септальный хрящ переходит в перпендикулярную пластину решетчатой кости, вывод, подтвержденный нашим исследованием.
Точная оценка хрящевой области полезна, поскольку она помогает в предоперационной оценке пациентов, перенесших ринопластику.Если хрящ, измеренный с помощью КТ, не подходит для запланированной операции, то оператор должен сообщить пациенту, что для забора хрящевого трансплантата необходим вторичный донорский участок. Еще одним преимуществом предоперационной оценки септального хряща является то, что она позволяет хирургу оценить, сколько трансплантата можно взять, сохраняя адекватную L-образную стойку сзади.
Использование информации о яркости для очерчивания септального хряща и расчета его размера является более сложным и может быть более точным по сравнению с методами, представленными в предыдущих исследованиях.Возможно, что значения, полученные с помощью нашего метода, более надежны, чем предыдущие измерения [1,5].
В нашем исследовании есть несколько ограничений, которые могли повлиять на результаты. Могут существовать различные отклонения, в том числе перекрытие между септальным хрящом и крыловидным хрящом или перекрытие трапециевидных областей, а также анатомические вариации. Чтобы свести к минимуму влияние этих факторов на результаты, авторы измерили расстояние между срезами КТ как 0.75 мм, что составляет примерно 1/3 от обычного измеренного значения. Цель этого состояла в том, чтобы свести к минимуму то количество перекрытий, которое происходит между структурами.
Даже если метод в нашем исследовании точен, невозможно было измерить все различия яркостей на всей границе между подкожной тканью и хрящевой тканью. Эту проблему можно решить, если использовать программу, которая может распознавать разницу в яркости между двумя тканями и точно устанавливать границу. В будущем можно будет точно измерить площадь хряща носовой перегородки перед операцией.Такой прогресс предоставит достаточно информации и данных не только пластическим хирургам, но и хирургам в других областях, таких как отоларингология, что в конечном итоге приведет к точной операции и хорошим результатам.
КТ-исследования могут быть очень полезными при предоперационной оценке хряща носовой перегородки. Используя разницу яркостей, мы смогли определить более точную границу хрящевой перегородки. КТ-исследования могут быть очень полезны при предоперационной оценке хряща носовой перегородки.Используя разницу яркостей, мы смогли определить более точную границу хрящевой перегородки. Чтобы этот метод был более автоматизированным и объективным, необходима дополнительная технология, которая может автоматически распознавать и указывать границу септального хряща. Таким образом, измерение хряща носовой перегородки с помощью КТ в будущем получит более широкое распространение, такой метод может предоставить важные данные для хирургии с использованием септального хряща, а также для предотвращения осложнений, таких как послеоперационные деформации носа.
Сноски
О потенциальном конфликте интересов, имеющем отношение к этой статье, не сообщалось.
Ссылки
1. Kim JH, Jung DJ, Kim HS, Kim CH, Kim TY. Анализ развития носовой перегородки и измерение септального хряща у корейцев с помощью трехмерной компьютерной томографии лицевых костей. Арх Пласт Хирург. 2014;41:163–170. [Бесплатная статья PMC] [PubMed] [Google Scholar]2. Саджадян А., Рубинштейн Р., Нагшине Н. Текущее состояние трансплантатов и имплантатов в ринопластике: часть I.аутологичные трансплантаты. Plast Reconstr Surg. 2010;125:40e–49e. [PubMed] [Google Scholar]3. Майлз Б.А., Петрисор Д., Као Х., Финн Р.А., Трокмортон Г.С. Анатомическая вариация носовой перегородки: анализ 57 трупных образцов. Отоларингол Head Neck Surg. 2007; 136: 362–368. [PubMed] [Google Scholar]4. Kim JS, Jang PY, Choi TH, Kim NG, Lee KS. Размер септального хряща при исследовании трупа. J Korean Soc Aesthetic Plast Surg. 2006; 12:29–32. [Google Академия]5. Ким И.С., Ли М.И., Ли КИ, Ким Х.И., Чон Ю.Дж.Анализ развития носовой перегородки в зависимости от возраста и пола с помощью МРТ. Clin Exp Оториноларингол. 2008; 1:29–34. [Бесплатная статья PMC] [PubMed] [Google Scholar]6. Ким Дж., Чо Дж. Х., Ким С. В., Ким Б. Г., Ли Д. С., Ким С. В. Анатомические вариации носовой перегородки: корреляция между компонентами перегородки. Клин Анат. 2010; 23:945–949. [PubMed] [Google Scholar]7. Homicz MR, McGowan KB, Lottman LM, Beh G, Sah RL, Watson D. Композиционный анализ хряща носовой перегородки человека. Arch Facial Plast Surg. 2003; 5:53–58.[PubMed] [Google Scholar]8. Коллаун С.С., Фикс Р.Дж., Мур М.Р., Васконез Л.О. Носовые хрящевые трансплантаты: более чем десятилетний опыт. Plast Reconstr Surg. 1997; 100:1547–1552. [PubMed] [Google Scholar]9. Тарди М.Э., младший, Деннени Дж., 3-й, Фрич М.Х. Универсальный хрящевой аутотрансплантат в реконструкции носа и лица. Ларингоскоп. 1985; 95: 523–533. [PubMed] [Google Scholar] 10. Гюнтер Дж. П., Рорих Р. Дж. Внешний доступ для вторичной ринопластики. Plast Reconstr Surg. 1987; 80: 161–174. [PubMed] [Google Scholar] 11.Веттер У., Хейт В., Хелбинг Г., Хайнце Э., Пирсиг В. Рост септального хряща человека: плотность клеток и образование колоний септальных хондроцитов. Ларингоскоп. 1984; 94: 1226–1229. [PubMed] [Google Scholar] 12. Ким Дже, Пак ХС, Юн Ч, Ким ХДж. Анализ перелома носовой перегородки в сочетании с переломом костей носа с помощью компьютерной томографии. J Korean Soc Plast Reconstr Surg. 1998; 25: 852–859. [Google Scholar]Операция по восстановлению перфорации носовой перегородки: процедура и восстановление
Вы смотрите на свою носовую перегородку каждый раз, когда смотрите в зеркало.Ну, во всяком случае, спереди. Это то, что отделяет вашу левую ноздрю от правой. И это проходит полностью внутри вашего носа. Он состоит из хряща спереди и кости сзади.
Если он перфорирован, это означает, что в нем есть отверстие. Это открывает путь от одной стороны вашего носа к другой.
Перфорация перегородки не всегда вызывает какие-либо симптомы, но они могут включать кровотечение из носа, затрудненное дыхание и ощущение заложенности носа. Вы можете издавать свистящие звуки при дыхании.
Примерно в половине случаев это происходит после операции по устранению другой проблемы с носом. Другие причины включают:
- Прижигание, лечение носовых кровотечений
- Некоторые заболевания, включая волчанку, некоторые виды рака, сифилис и туберкулез
- Повреждение носа, например, его поломку или даже чрезмерное ковыряние
- Инфекции
- Использование лекарств, которые слишком часто попадают в нос, от безрецептурных назальных спреев до кокаина
Иногда хирургическое вмешательство может исправить перфорированную перегородку, но оно не всегда работает и не всегда необходимо.Как правило, маленькие отверстия легче исправить, чем большие.
Что происходит во время операции?
Процедура занимает от 1 до 3 часов, и обычно вы уходите домой в тот же день. Обычно вы принимаете наркотики, чтобы помочь вам проспать это.
Некоторые врачи используют закрытую хирургию, что означает, что они входят через отверстие в ваших ноздрях. Другие открывают нос для лучшего доступа. Есть всевозможные способы исправления. Ваш врач выберет один из них в зависимости от того, где находится отверстие и насколько оно велико, а также в зависимости от своих навыков и опыта.
Как правило, они берут ткань из вашего носа или даже изо рта, чтобы закрыть отверстие. Затем они помещают лоскут новой ткани с каждой стороны отверстия.
Некоторые врачи считают, что это все, что вам нужно, в то время как другие добавляют дополнительную поддержку кости, хряща или ткани из других частей тела.
Что такое восстановление?
У вас, вероятно, будут тонкие шины в носу, чтобы покрыть и защитить закрылки. Они останутся на неделю или две.
Чтобы защитить нос во время его заживления, лучше всего:
- Не поднимать тяжелые предметы и не наклоняться
- Не ковырять в носу и не сморкаться слишком сильно
- Промывать физиологическим раствором так часто, как вам скажет врач
- Чихайте с открытым ртом
- Держитесь подальше от контактных видов спорта
В большинстве случаев вы почувствуете себя нормально в течение нескольких недель.
Вам, вероятно, предстоит контрольный визит через неделю после операции. Ваш врач:
- Проверит шины и может снять их
- Прочистит нос, если у вас есть корки
- Убедится, что вы хорошо заживаете 6 и 12 недель. Обычно вы знаете, сработало ли это после 6 недель.
Когда следует звонить врачу?
Как и при любой операции, во время заживления нужно внимательно следить за состоянием.Позвоните своему врачу, если у вас есть:
- Кровотечение
- Лихорадка
- Обильное выделение жидкости из носа
- Боль и отек, которые усиливаются
- Покраснение на внешней стороне носа
- Проблемы с дыханием 72 72 9 Знайте, нужна ли мне операция в первую очередь?
Хирургия обычно не является первым шагом в лечении. Ваш врач может предложить мазь или обычное полоскание солевым раствором. Если это облегчит ваши симптомы, вам все равно придется проходить регулярные осмотры, потому что дыра может расти.Если он станет слишком большим, ваш нос не будет таким стабильным, и это может привести к тому, что кончик или середина вашего носа начнут опускаться.
Если они не работают, ваш врач может искать другие проблемы с вашим носом, особенно если ваш основной симптом заключается в том, что вы чувствуете закупорку. Это может быть вызвано чем-то другим, и операция может не помочь.
Также нужно понять, что привело к дыре. Если это было вызвано постоянной проблемой, например опухолью, инфекцией или употреблением наркотиков, ваш врач должен лечить ее в первую очередь.Тогда можно подумать об уходе за перегородкой.
Медицинская школа Восточной Вирджинии (EVMS), Норфолк, Хэмптон Роудс
EVMS Хирурги уха, горла и носа
Синус
Септопластика – операция по выпрямлению носовой перегородки. На этой странице более подробно описано, когда рекомендуется эта операция, как она проводится и чего ожидать, если вам предстоит септопластика.
Носовая перегородка — это часть носа, которая отделяет правую носовую полость от левой носовой полости. Перегородка обычно лежит прямо в центре носа и опирается на костный гребень, называемый верхнечелюстным гребнем. Перегородка состоит из нескольких слоев. Спереди центральная часть перегородки представляет собой хрящ, относительно мягкий и достаточно гибкий материал (на диаграмме окрашен синим цветом). Сзади центральная перегородка состоит из тонкой кости (светло-коричневая на диаграмме).Поверх костей и хрящей лежит слизистая оболочка носа, называемая слизистой оболочкой.
На рисунке слева показана внутренняя структура носовой перегородки в разрезе. Передняя часть носа слева. Заштрихованная область состоит из хряща (синий). В задней части перегородка состоит из тонкой кости (светло-коричневого цвета).
Септопластика проводится, если перегородка искривлена настолько, что мешает дыханию или вызывает другие проблемы.Немногие люди на самом деле имеют идеально прямую перегородку, но в большинстве случаев степень изгиба недостаточно велика, чтобы ее можно было заметить.
На этом изображении показан нос, вид снизу. В этом случае перегородка отклонена в правую сторону больного. В некоторых случаях передняя часть перегородки находится в правильном положении, а задняя часть имеет отклонение. Наиболее распространенным симптомом сильно искривленной (кривой) перегородки является затрудненное дыхание через нос. Симптомы обычно усиливаются на одной стороне, а иногда и на стороне, противоположной изгибу.В некоторых случаях искривленная перегородка может мешать дренажу пазух, что приводит к повторным инфекциям пазух. Также может потребоваться выпрямление перегородки у лиц, перенесших операцию на носовых пазухах, только для того, чтобы инструменты, необходимые для этой операции, можно было поместить в носовую полость.
Другие причины заложенности носа
Важно помнить, что нос проходит через естественный цикл, в котором линия носа сначала с одной, а затем с другой стороны медленно расширяется и сужается.Эти циклы происходят каждые несколько часов и могут привести к ощущению заложенности носа. Если у вас постоянно затруднено дыхание через нос, вам следует обратиться к отоларингологу для полного обследования. Вы НЕ должны использовать какие-либо безрецептурные назальные спреи в течение более чем нескольких дней, не связавшись с врачом. Многие из этих продуктов содержат ингредиенты, которые вызывают отек носа, когда они прекращаются. В результате пользователь должен продолжать использовать спрей, чтобы сохранить чистоту своего носа.Такое длительное использование может привести к повреждению слизистой оболочки носа. Есть спреи, которые можно использовать длительное время без побочных эффектов; спросите своего врача, какой спрей лучше всего подходит для вас.
Септопластика: выполнение операции
Основным принципом септопластики является выпрямление искривленных участков перегородки. Операция почти всегда делается через нос, без наружного разреза (разрез может быть, если одновременно делается и ринопластика).Сильно искривленные участки перегородки могут быть удалены полностью или же они могут быть удалены, отрегулированы и повторно вставлены в нос. Операция занимает около 1-1,5 часов, в зависимости от сложности отклонения. Это может быть сделано под местной или общей анестезией и обычно проводится амбулаторно. Во многих случаях назальные тампоны помещаются на срок от одного до нескольких дней. Почти никогда не бывает синяков на коже только из-за септопластики, хотя синяки могут быть, если одновременно проводится ринопластика.
Возможные осложнения септопластики
Если операция проводится под общей анестезией, существуют обычные риски при любой общей анестезии. Кровотечение является возможным риском, который может потребовать тампонирования носа. Гораздо реже риск образования отверстия в перегородке после операции. Это может вызвать корки в носу или помешать дыханию через нос. Еще более редким осложнением является послеоперационная инфекция перегородки, которая может привести к искривлению передней части носа внутрь.Это потребовало бы второй операции, чтобы исправить деформацию.
Как узнать, нужна ли мне септопластика?
Лучше всего пройти осмотр носа у отоларинголога. Заложенность носа может быть вызвана не только искривлением носовой перегородки, но и другими причинами, поэтому важно провести тщательное обследование носа. заболевание, характеризующееся утратой хрящевых и/или костных структур носовой перегородки вместе с их слизисто-надхрящничной и слизисто-надкостничной оболочками.Существует много причин для NSP, таких как хирургия носа в анамнезе (ятрогенная), травма, самоповреждения, прием лекарств, химические раздражители, неопластические причины, воспалительные причины (васкулиты, гранулематоз Вегенера) или инфекции (сифилис, туберкулез). Перфорация носовой перегородки создает патологические завихрения в носовом потоке воздуха, что приводит к снижению нормального процесса увлажнения. В зависимости от размера и локализации дефекта это может привести к серьезному дискомфорту пациентов и снижению качества их жизни.
Поскольку современные методы лечения (использование синтетического материала, PDS или нативного хряща) имеют недостатки, исследователи хотят показать в исследовании фазы I, что использование тканевой инженерии аутологичного носового хряща для восстановления перфораций (отверстий) в этой области носовой перегородки подходит и безопасен. Сконструированный аутологичный трансплантат носового хряща используется в качестве замены PDS или нативного хряща в промежуточном трансплантате при реконструкции перфораций носовой перегородки.
Перед операцией регистрируют субъективные симптомы пациентов с помощью двух опросников: Оценки симптомов назальной обструкции (NOSE) и визуальной аналоговой шкалы (ВАШ) и проводят объективные исследования с функциональными тестами (риноманометрия, акустическая ринометрия) и визуализацией (эндоскопия). , КТ). Небольшой биоптат, взятый из носовой перегородки, обрабатывается в лаборатории, хондроциты выделяются, расширяются и культивируются на коллагеновой матрице для создания тканеинженерного хрящевого трансплантата.