Изменение формы носа в Москве от 160 000 руб
Эффект ринопластики
2 недели после операции вам придется носить фиксирующую повязку. Также на весь последующий месяц нужно отказаться от острых, слишком горячих или слишком холодных блюд, интенсивных физических нагрузок и ношения очков. Постарайтесь по минимуму напрягать мимические мышцы и будьте особенно осторожны, если снимаете одежду через голову.
После реабилитационного периода ваш нос приобретет именно ту форму, которую вы желали видеть. Он станет более пропорциональным, с четко очерченной переносицей, аккуратными ноздрями, крыльями и кончиком.
Записаться на прием квалифицированного хирурга «СМ-Пластика», узнать детали и стоимость процедуры можно, заполнив форму обратной связи на сайте или позвонив по телефону в Москве: +7 (495) 777-48-05.
Пластическая хирургия не остается в стороне от современных тенденций в медицине, а именно перехода к минимально-инвазивным хирургическим вмешательствам.
Использование классических методов остеотомии при проведении ринопластики с применением долот и молотка уходит в прошлое. Наиболее важным в остеотомии является четкое соблюдение уровня рассечения кости с получением ровного среза и, как следствие, предупреждение развития грубой костной мозоли в будущем. В случае применения долот риск неконтролируемой остетомии усугубляется, опасность оскольчатого перелома носовых костей при остеопорозе увеличивается, кроме того в связи с травматизацией мягких тканей в течение 3-4 недель сохраняется отек и гематомы.
Незаменимым помощником в случае проведения ринопластики с остеотомией стало применение пьезоинструмента. Помимо более качественной остетомии (сохраняющей структуру спинки носа) пьезоинструмент обладает ещё одним преимуществом – это сохранение мягкотканных структур, в том числе, и сосудов в зоне остеотомии.
Таким образом, неоспоримыми преимуществами метода являются:
- Минимальная травма мягких тканей.
- Очень точная и предсказуемая работа на кости.
- Дополнительные асептика и гемостаз во время операции.
Изменение формы носа — сделать пластику носа в клинике СОЮЗ
Техника изменения формы носа
Перед вмешательством нужно будет пройти детальное обследование. Затем с помощью компьютерного моделирования подбирается форма носа, идеально подходящая конкретному пациенту. Перед тем, как
 На консультации также определяется цена пластики носа. После решения всех вопросов, и получения данных обследования, пациент направляется на коррекцию.
На консультации также определяется цена пластики носа. После решения всех вопросов, и получения данных обследования, пациент направляется на коррекцию.Операция выполняется под местным обезболиванием с дополнительной седацией либо под общим наркозом. Вмешательство длится от одного до трех часов, это зависит от сложности и объема пластики.
Различают два вида ринопластики:
- Закрытая – все разрезы осуществляются внутри носовых ходов, поэтому после процедуры не остается заметных шрамов и короче период восстановления. В тоже время закрытый вариант операции намного сложнее открытого и требует от хирурга большого опыта;
- Открытая – к ней прибегают, если нужны обширные манипуляции на костной и хрящевой ткани, которые требуют дополнительного открытого доступа. В этом случае разрез производится на колумелле – срединной части носа, в результате чего все его хрящи открываются и становятся доступными для действий хирурга.
После коррекции на область носа накладывается гипсовая лангета, а в носовые ходы вставляются тампоны. После операции показано назначение антибиотиков и поддерживающей терапии. Для укрепления иммунитета применяют иммуномодулирующие средства. Полное восстановление поврежденных тканей происходит примерно через 3-4 месяца. Однако изменения формы носа заметны уже через две недели. Пациент во время восстановительного периода должен избегать сильных физических нагрузок, посещения сауны и бани, употребления алкогольных напитков. Даже после окончательного восстановления прооперированный нос нужно беречь от травм, а также стараться избегать развития хронических воспалительных заболеваний – ринитов и синуситов.
После операции показано назначение антибиотиков и поддерживающей терапии. Для укрепления иммунитета применяют иммуномодулирующие средства. Полное восстановление поврежденных тканей происходит примерно через 3-4 месяца. Однако изменения формы носа заметны уже через две недели. Пациент во время восстановительного периода должен избегать сильных физических нагрузок, посещения сауны и бани, употребления алкогольных напитков. Даже после окончательного восстановления прооперированный нос нужно беречь от травм, а также стараться избегать развития хронических воспалительных заболеваний – ринитов и синуситов.
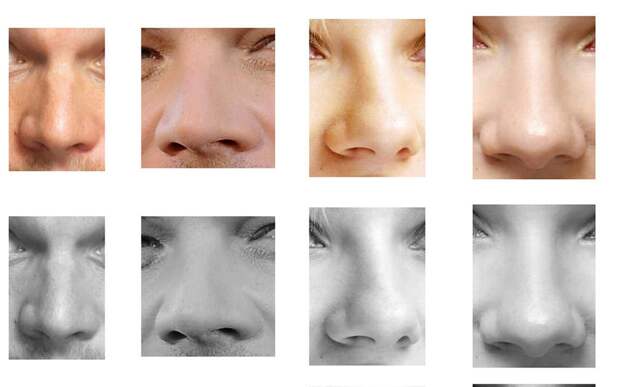
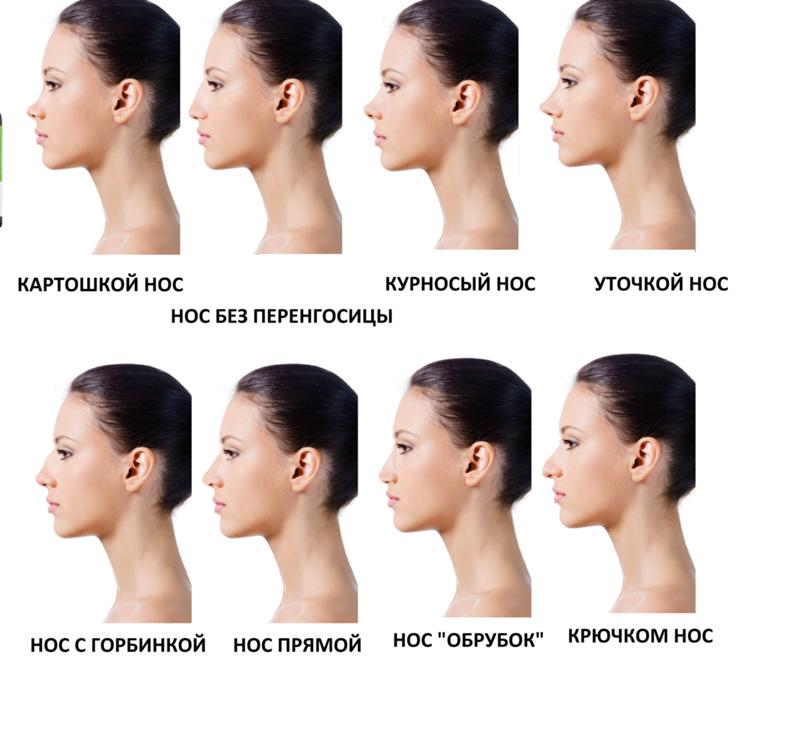
Форма носа | Prof. Dr. Berghaus
Форма носа определяется, главным образом, хрящевой и костной структурой. Кроме того, к формообразованию носа причастны мягкие ткани и кожный покров. При этом различают безвредные, но при определенных обстоятельствах серьезные для пациента аномалии внешнего формообразования носа, такие как горбинка, изогнутость или сколиоз носа, а также слишком широкий или седловидный нос. В дополнение к этим частым изменениям могут иметь место настоящие дефекты развития, такие как деформация носа при полной расщелине губы и нёба. Кроме того, в результате опухолей или несчастных случаев могут возникать явные дефекты вплоть до полной потери носа.
В дополнение к этим частым изменениям могут иметь место настоящие дефекты развития, такие как деформация носа при полной расщелине губы и нёба. Кроме того, в результате опухолей или несчастных случаев могут возникать явные дефекты вплоть до полной потери носа.
Коррекция носа в Мюнхене
Солидные знания в области анатомии, обширный опыт операций и использование современных, передовых хирургических методов, а также точные хирургические инструменты, позволяют исправить большинство аномалий формы и дефектов развития носа, вплоть до полноценной хирургической замены (в случае полного отсутствия носа).
Хирургические методы коррекции носа
При необходимости, коррекция носа в Мюнхене проводится профессором Berghaus вместе с операцией по устранению функциональных нарушений во внутренней области носа, препятствующих нормальному носовому дыханию. Для коррекции внешней формы носа, т.е. ринопластики, в принципе, существуют, так называемые, «открытые» и «закрытые» (эндоназальные) хирургические методы.
Открытая коррекция носа
При открытых методах делают небольшой разрез на переносице с тем, чтобы можно было освободить от кожи хрящевую и костную структуру и, таким образом, сделать ее более обозримой и доступной. Однако такой разрез несет с собой определенный риск образования рубцов и деформаций на переносице, что, впрочем, случается очень редко. При использовании надлежащей хирургической техники, бинокулярной лупы и тонкого как волос материала швов открытые методы оставляют, как правило, едва заметные рубцы.
Закрытая коррекция носа
Для того чтобы сохранить в неприкосновенности весь кожный покров носа и избежать образования рубцов на переносице, в соответствующих случаях может быть выбран интраназальный (т.н. «закрытый») хирургический метод. При соответствующем опыте можно также и с помощью этого щадящего метода целенаправленно обеспечить различные изменения формы носа. В зависимости от необходимости, а также от желания пациента, в отдельных случаях изменения формы носы выполняют резекцию, фиксацию швов или принимают меры для усиления структуры носа.
Для выпрямления или сужения формы в обоих методах, при необходимости, может потребоваться целенаправленное расчленение носовой кости (остеотомия).
Для оптимизации результатов, операция в Мюнхене делается профессором Berghaus с использованием специальных хирургических очков с сильным увеличением. При этом для швов используется преимущественно очень тонкий материал, который в большинстве случаев растворяется в течение короткого времени, так что большая часть ниток устраняется, и в мягких тканях, как правило, не остается никаких остатков.
Перед коррекцией носа
Для подготовки операции в Мюнхене требуется точный анализ медицинского заключения, который проводится профессором Berghaus вместе с пациентом. При планировании операции учитываются пожелания и идеи пациентов и охотно привлекаются обработанные на компьютере изображения предполагаемой формы носа (т.н. ‚Симуляция‘; см. „Цифровое ателье носа“). Операции коррекции формы носа у профессора Berghaus в целом могут быть скомбинированы с операциями на носовой перегородке, носовой раковине, околоносовых пазухах и т.
После коррекции носа
После коррекции внешней формы носа в Мюнхене, для стабилизации на спинке носа на период 12 дней устанавливается алюминиевая шина. В первые дни, по мере необходимости, используются охлаждающие накладки (подушечки) для лица, чтобы предотвратить отек мягких тканей.
Для внутренней опоры носа и облегчения дыхания, в течение трех дней носят в носу мягкие силиконовые трубки, которые не причиняют беспокойства и очень легко удаляются. К тампонированию, которое затрудняет носовое дыхание, прибегают лишь в очень редких случаях.
После коррекции носовой перегородки и внешней формы носа (септоринопластика), возможные отеки и проявления дисхромии в большинстве случаев практически полностью исчезают после окончательного удаления внешней носовой шины, т.е. примерно через две недели. В результате постепенного исчезновения отека кожи на протяжении нескольких месяцев новая форма носа становится более четкой.
Вас могут заинтересовать также следующие темы:
Коррекция формы носа в Германии, стоимость лечения
Операционная коррекция формы носа помогает изменить его размер, форму переносицы или исправить форму его кончика.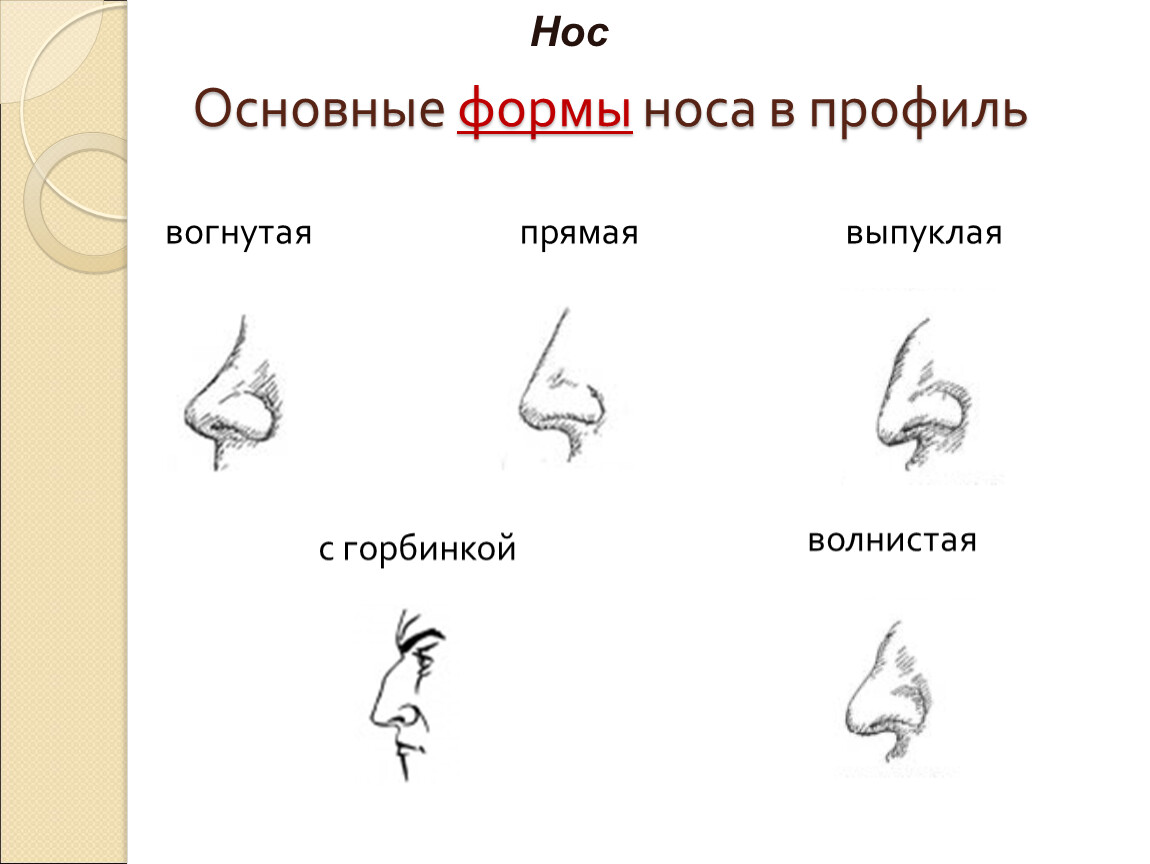
Различают эстетическую и функциональную ринопластику.
Ринопластика позволяет исправить, как эстетическое недовольство формой носа, так и его функциональные проблемы.
Цель эстетической коррекции формы носа в Германии – создание наиболее гармоничной формы носа, идеально подходящей к Вашему лицу.
Основные показания к операции по коррекции формы носа:
- Изменение формы носа на основе пожеланий клиента
- Устранение врожденного анатомического дефекта
- Исправление травматических повреждений
- До 20 лет формируются кости лицевого черепа, поэтому до достижения этого возраста ринопластика считается противопоказанной. Исключением являются серьезные медицинские показания или врожденные пороки.
- После 40 лет стараются избегать оперативного вмешательства из – за ослабления регенеративных процессов и большего риска осложнений.
- Сердечно — сосудистые заболевания в стадии неполной компенсации.

- Сахарный диабет первого и второго типа в стадии неполной компенсации.
- Нарушение свертываемости крови
- Онкологические заболевания
- Острые вирусные заболевания
- Воспаление кожи на лице
- Патология печени и почек в стадии неполной компенсации.

Перед операцией по коррекции формы носа в пластической клинике Германии Вам будет предоставлена индивидуальная консультация с Вашим лечащим врачом – пластическим хирургом — для детального исследования лица и подбора оптимальной методики коррекции формы носа на основании Ваших пожеланий и медицинских показаний.
В зависимости от индивидуальных факторов (Ваш возраст, структура костей и хрящей носа, толщина кожи и личные пожелания), определяется объем и метод проведения операции.
Метод проведения ринопластики:
Высокоспециализированный хирург пластической клиники в Германии, по — новому формирует костно-хрящевую основу, делая разрез в области переносицы (открытая ринопластика) приподнимая кожу. В случае коррекции носа с горбинкой, может быть удалены излишние костная или хрящевая ткани.
В случае коррекции носа с горбинкой, может быть удалены излишние костная или хрящевая ткани.
При закрытой ринопластике хирургический доступ к структурам хуже, но также эффективен. Во время закрытой ринопластики разрезы делают в самой полости носа, обеспечивая необходимую форму.
После операции в носовые пазухи устанавливают специальные тампоны, которые останавливают кровотечение и поддерживают правильную форму носа. Затем накладывают пластырную повязку и шину, как компрессионное средство для предотвращения кровоизлияния и отеков.
Продолжительность операции по коррекции формы носа в Германии:
От 1,5 до 2,5 часов.
Обезболивание:
Ринопластика в Германии проводится в основном под полным наркозом, в некоторых случаях — с применением местной анестезии.
Реабилитация
Через 7 – 8 дней снимают повязку с носа, в это время виден предварительный результат операции. Небольшие отеки могут сохраняться в течение нескольких месяцев. Вопрос о времени перелета домой решается индивидуально с лечащем врачом. Окончательные изменения очевидны после нескольких месяцев заживления и восстановления.
Окончательные изменения очевидны после нескольких месяцев заживления и восстановления.
После ринопластики:
- На два месяца запрещено заниматься спортом, посещать бассейн и загорать
- Три недели необходимо есть и пить только теплую еду и напитки
- нежелательно употребление алкоголя и курение — это препятствует заживлению
Весь период реабилитации после проведения коррекции формы носа в Германии Вы можете провести в экологически чистой зоне в условиях 4* отеля Stadt Freiburg, на последнем этаже которого расположена клиника пластической хирургии имени Эриха Лексера, что сочетает в себе получение высококлассных медицинских услуг и гарантирует Вам комфортное пребывание. Пациенты могут принимать пищу, не выходя из номера, а также пользоваться специальным лифтом, который соединяет их палаты с кабинетами лечащих врачей.
Мы гарантируем индивидуальный подход к каждому клиенту и полную конфиденциальность процесса лечения. После проведения ринопластики в Германии Вы сможете оценить отменный результат, который будет радовать Вас долгие годы.
Хирург рассказал, может ли медицинская маска изменить форму носа и ушей
2021-11-21T01:13:00+03:00
2021-11-21T08:13:30+03:00
2021-11-21T01:13:00+03:00
2021
https://1prime.ru/health/20211121/835283515.html
Хирург рассказал, может ли медицинская маска изменить форму носа и ушей
Здоровье
Новости
ru-RU
https://1prime.ru/docs/terms/terms_of_use.html
https://россиясегодня.рф
Плотно прилегающая к лицу медицинская маска может давить на нос, и некоторые граждане жалуются, что нос «сплющился» или даже «опустился вниз». Пластический хирург Гайк Бабаян в… ПРАЙМ, 21.11.2021
общество , здоровье, новости, экспертное мнение, сиз, covid — 19, россия
https://cdnn.1prime.ru/images/83307/24/833072410.jpg
1920
1440
true
https://cdnn.1prime.ru/images/83307/24/833072410.jpg
https://cdnn.1prime.ru/images/83307/24/833072409.jpg
1920
1080
true
https://cdnn. 1prime.ru/images/83307/24/833072409.jpg
1prime.ru/images/83307/24/833072409.jpg
https://cdnn.1prime.ru/images/83307/23/833072397.jpg
1920
1920
true
https://cdnn.1prime.ru/images/83307/23/833072397.jpg
https://1prime.ru/health/20211120/835280567.html
Агентство экономической информации ПРАЙМ
7 495 645-37-00
ФГУП МИА «Россия сегодня»
https://россиясегодня.рф/awards/
Агентство экономической информации ПРАЙМ
7 495 645-37-00
ФГУП МИА «Россия сегодня»
https://россиясегодня.рф/awards/
Агентство экономической информации ПРАЙМ
7 495 645-37-00
ФГУП МИА «Россия сегодня»
https://россиясегодня.рф/awards/
Агентство экономической информации ПРАЙМ
7 495 645-37-00
ФГУП МИА «Россия сегодня»
https://россиясегодня.рф/awards/
Агентство экономической информации ПРАЙМ
7 495 645-37-00
ФГУП МИА «Россия сегодня»
https://россиясегодня.рф/awards/
МОСКВА, 21 ноя — ПРАЙМ. Плотно прилегающая к лицу медицинская маска может давить на нос, и некоторые граждане жалуются, что нос «сплющился» или даже «опустился вниз». Пластический хирург Гайк Бабаян в беседе с радио Sputnik заявил, что беспокоиться по этому поводу не стоит.
Плотно прилегающая к лицу медицинская маска может давить на нос, и некоторые граждане жалуются, что нос «сплющился» или даже «опустился вниз». Пластический хирург Гайк Бабаян в беседе с радио Sputnik заявил, что беспокоиться по этому поводу не стоит.
Вирусолог спрогнозировал, когда в России спадет волна COVID-19
По его словам, от ношения медицинской маски нос действительно может опуститься, но это изменение длится 10-15 минут после снятия маски, а потом нос «со стопроцентной вероятностью вернется обратно». «Если быть объективным, то маска никак на форму лица или отдельных частей лица не может влиять. Без каких-либо дополнительных вмешательств все восстанавливается, природа так создала», — заверил Бабаян.
Также он отметил, что резинки медицинской маски могут давить на уши — но и это не способно привести к каким-лидо последствиям.
«Ушные раковины очень эластичны и у них сильна тканевая память формы. Под влиянием внешних факторов ушные раковины не могут измениться. Это исключено. Даже больше скажу, после хирургического вмешательства уши могут возвращать свою прежнюю форму и приходится делать еще одну коррекцию», — объяснил пластический хирург.
Это исключено. Даже больше скажу, после хирургического вмешательства уши могут возвращать свою прежнюю форму и приходится делать еще одну коррекцию», — объяснил пластический хирург.
Ринопластика (коррекция формы носа)
Цель операции
В медицинском центре Шамир — «Ассаф Арофе» проводятся множество операций по коррекции формы носа. Операции проводятся опытным и профессиональным персоналом после нескольких предварительных встреч с пациентом. Вид операции зависит от ее цели. Квалификация медицинского персонала в отделении позволяет проводить широкий спектр операций по коррекции носа, таких как:
- Уменьшение носа или его реконструкция посредством имплантатов
- Удаление горбинки
- Изменение угла между носом и губой
- Коррекция ямки в области корня носа
- Коррекция носовой перегородки
- Уменьшение носовых раковин
Необходимо подчеркнуть, что подобную операцию можно сочетать с процедурами для лечения проблем с дыханием.
Подходящие кандидаты
Наилучшим кандидатом является пациент, желающий улучшить свой внешний вид, но не стремящийся к совершенству. Если клиент является физически здоровым, психологически стабильным и с реальными ожиданиями — он подходит для прохождения операции.
Сама операция
Операция может проводиться под общей или местной анестезией. Операция проводится через ноздри, и в ее ходе врач корректирует кости и хрящи носа, не трогая саму кожу. После операции пациент будет страдать от отека в области носа, верхней губы и верхних и нижних век. Возможны также подкожные кровоизлияния, особенно вокруг глаз. Все они пройдут через несколько дней/недель.
Планирование операции
Планирование операции играет значительную роль в ее успехе. На первичной консультации хирурга спросит пациента, какую форму носа он хотел бы иметь. Хирург осмотрит нос изнутри и извне, изучит пропорции носа относительно лица и обсудит различные варианты лечения. Хирург объяснит, какие факторы влияют на конечный результат операции: строение костей и хрящей носа, черты лица, толщина кожи и возраст пациента.
Пациент должен сообщить хирургу о любых операциях на носу, перенесенных в прошлом, о любых травмах носа, даже очень старых, о насморке, аллергиях или других проблемах с дыханием. Также хирургу необходимо сообщить о фоновых заболеваниях, аллергиях на лекарства, постоянно принимаемых лекарствах, привычках курения и употребления алкоголя и наркотиков.
Коррекция формы носа — Медицинский центр «ЗДОРОВЬЕ»
Коррекция искривленного носа в Брянске
В медицинском центре «Здоровье» можно исправить любые искривления носа с помощью современных методов пластической хирургии. Как центральная часть, нос оказывает сильное влияние на восприятие лица окружающими. Конечно же, любые искривления носа не останутся незамеченными. Это нередко становится поводом для ринопластики. Однако, эстетический вопрос не является единственной причиной пластики искривленного носа.
Показаниями к ринопластике и септопластике (пластика перегородки носа) могут стать нарушения дыхания и хронические заболевания в области носоглотки, такие как риниты, синуситы, носовые кровотечения. Проблемы в этой области также могут стать причиной постоянных головных болей.
Проблемы в этой области также могут стать причиной постоянных головных болей.
Искривления носа бывают простые, когда происходит смещение от центра лица в ту или иную сторону или более сложные изогнутой формы. Такие дефекты возникают из-за:
— врожденных или приобретенных травм,
— проблем в раннем детском возрасте — перенесенных заболеваний, таких как рахит;
— неравномерного роста костей.
Принято выделять два типа искривлений носа – хрящевые и костные. Асимметрия носа, как правило, проявляется не только по спинке носа, но и в других частях, поэтому коррекция может затрагивать кончик носа, его крылья и обязательно перегородку.
Коррекция искривлений носа в медицинском центре «Здоровье» в Брянске
Чтобы избавиться от проблемы искривленного носа нужно прежде всего записаться на консультацию к специалисту. Можно долго выбирать и читать отзывы о врачах, но все же лучше прийти и пообщаться с доктором лично. Попросить его показать фото результатов работы и рассказать, что можно сделать в вашем случае.
Если работа пластического хирурга вам понравилась и вы нашли общий язык с доктором, вы можете получить направление на обследование и анализы. Это очень важный этап, который позволяет оценить состояние вашего здоровья, исключить случаи противопоказаний и риски побочных эффектов. Обычно требуются анализы крови, мочи, заключение терапевта и анестезиолога. Кроме этого, необходимы результаты рентгенографии костей носа и заключение оториноларинголога.
Предварительно оценить эффективность операции в вашем случае можно с помощью компьютерного моделирования. Все анализы и обследования можно пройти в короткие сроки и без очередей в нашем медицинском центре.
Подготовка к операции коррекции кривого носа
Доктор на консультации подробно расскажет как правильно подготовиться к операции в вашем случае, чтобы реабилитация прошла быстро и успешно.
Ход операции
Операция по коррекции носа выполняется под общим наркозом и длится около двух часов. Чтобы придать носу правильную форму доктор делает разрезы на слизистой и перемещает ткани, удаляя лишние части, чтобы костный и хрящевой отделы приобрели правильные очертания. После этого накладываются швы, а для удержания формы специальная лонгета. Для фиксации формы, предотвращения смещения тканей и остановки кровотечения, в ноздри устанавливают турунды – марлевые тампоны. При необходимости может быть проведена контурная пластика гиалуроновой кислотой специальной консистенции.
После этого накладываются швы, а для удержания формы специальная лонгета. Для фиксации формы, предотвращения смещения тканей и остановки кровотечения, в ноздри устанавливают турунды – марлевые тампоны. При необходимости может быть проведена контурная пластика гиалуроновой кислотой специальной консистенции.
После операции врач назначает обезболивающие препараты и антибиотики, которые подбираются индивидуально. Период реабилитации длится несколько месяцев. Через 3-5 дней после операции убирают турунды, которые доставляют основное неудобство пациенту.
В первые дни после операции нельзя дышать носом, рекомендуется придерживаться постельного режима, не употреблять слишком горячую или холодную пищу и напитки. Не рекомендуются физические нагрузки. Обычно швы снимают на десятый день. Не рекомендуются физические нагрузки в течение месяца. Предварительный результат можно оценить когда спадут отеки и синяки. Окончательное заключение об эффективности операции можно сделать не ранее чем, через полгода.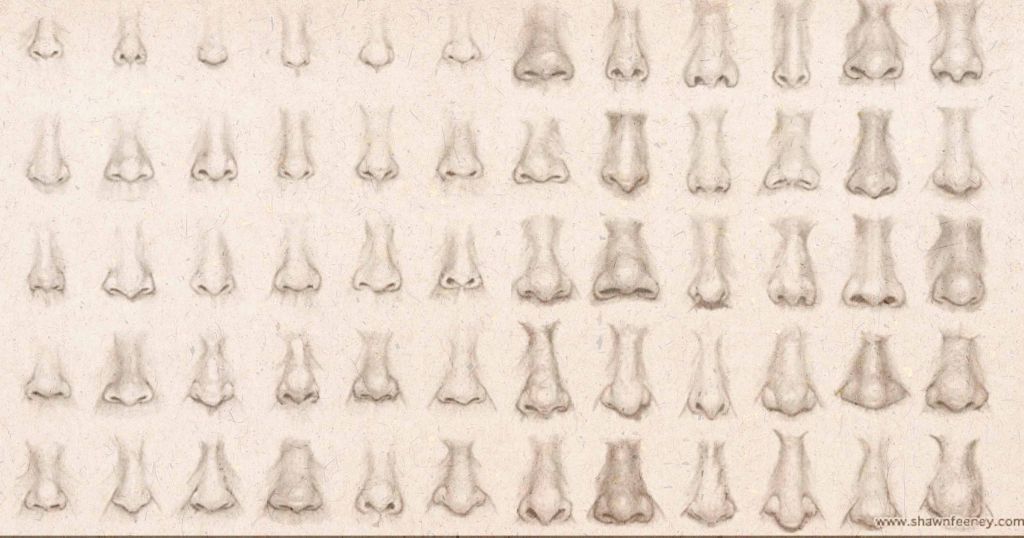
Узнать подробнее о проведении коррекции кривого носа, цене и других нюансах можно позвонив администратору и записавшись на консультацию. А также можно заполнить онлайн форму на нашем сайте.
Расследование случая формы человеческого носа и адаптации к климату
Abstract
Эволюционные причины вариаций формы носа в разных популяциях людей постоянно обсуждаются. Важной функцией носа и носовой полости является кондиционирование вдыхаемого воздуха до того, как он достигнет нижних дыхательных путей. По этой причине считается, что наблюдаемые различия в форме носа среди популяций не просто результат генетического дрейфа, но могут быть адаптацией к климату.Чтобы ответить на вопрос, является ли местная адаптация к климату ответственной за дивергенцию формы носа у популяций, мы используем сравнения Qst – Fst, чтобы показать, что ширина ноздрей и ширина крылового основания более дифференцированы в популяциях, чем ожидалось при одном только генетическом дрейфе. Чтобы проверить, связана ли эта дифференциация с адаптацией к климату, мы сравнили пространственное распределение этих переменных с глобальным распределением температуры, абсолютной влажности и относительной влажности. Мы обнаружили, что ширина ноздрей коррелирует с температурой и абсолютной влажностью, но не с относительной влажностью.Мы пришли к выводу, что некоторые аспекты формы носа действительно могли быть обусловлены местной адаптацией к климату. Однако мы думаем, что это упрощенное объяснение очень сложной эволюционной истории, которая, возможно, также включала другие ненейтральные силы, такие как половой отбор.
Чтобы проверить, связана ли эта дифференциация с адаптацией к климату, мы сравнили пространственное распределение этих переменных с глобальным распределением температуры, абсолютной влажности и относительной влажности. Мы обнаружили, что ширина ноздрей коррелирует с температурой и абсолютной влажностью, но не с относительной влажностью.Мы пришли к выводу, что некоторые аспекты формы носа действительно могли быть обусловлены местной адаптацией к климату. Однако мы думаем, что это упрощенное объяснение очень сложной эволюционной истории, которая, возможно, также включала другие ненейтральные силы, такие как половой отбор.
Информация об авторе
Изучение адаптации человека имеет важное значение для нашего понимания этиологии болезни. Эволюционные исследования того, почему определенные фенотипы заболеваний, такие как серповидноклеточная анемия и непереносимость лактозы, возникают с разной скоростью в разных популяциях, привели к лучшему пониманию вовлеченных генетических и экологических факторов риска. Точно так же исследования географического распределения пигментации кожи продолжают давать важные подсказки относительно риска дефицита витамина D и рака кожи. Здесь мы исследуем, было ли изменение формы носа у разных популяций вызвано региональными различиями в климате. Мы обнаружили, что вариации как в ширине ноздрей, так и в ширине крылового основания, по-видимому, ускоренно разошлись среди человеческих популяций. Мы также обнаружили, что геопространственное распределение ширины ноздрей коррелирует с температурой и абсолютной влажностью, но не с относительной влажностью.Наши результаты подтверждают утверждение о том, что местная адаптация к климату могла сыграть роль в эволюции различий формы носа в разных популяциях людей.
Точно так же исследования географического распределения пигментации кожи продолжают давать важные подсказки относительно риска дефицита витамина D и рака кожи. Здесь мы исследуем, было ли изменение формы носа у разных популяций вызвано региональными различиями в климате. Мы обнаружили, что вариации как в ширине ноздрей, так и в ширине крылового основания, по-видимому, ускоренно разошлись среди человеческих популяций. Мы также обнаружили, что геопространственное распределение ширины ноздрей коррелирует с температурой и абсолютной влажностью, но не с относительной влажностью.Наши результаты подтверждают утверждение о том, что местная адаптация к климату могла сыграть роль в эволюции различий формы носа в разных популяциях людей.
Образец цитирования: Zaidi AA, Mattern BC, Claes P, McEcoy B., Hughes C, Shriver MD (2017) Расследование случая формы человеческого носа и адаптации к климату. PLoS Genet 13 (3):
e1006616.
https://doi.org/10. 1371/journal.pgen.1006616
1371/journal.pgen.1006616
Редактор: Грег Гибсон, Технологический институт Джорджии, США
Поступила: 17 декабря 2015 г .; Одобрена: 3 февраля 2017 г .; Опубликован: 16 марта 2017 г.
Авторские права: © 2017 Zaidi et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Мы сделали почти все данные, которые использовались для получения результатов в документе, общедоступными в электронной таблице в разделе вспомогательной информации (файл S1). Таблица включает обезличенные данные, разделенные по анализам, для которых они использовались.В частности, мы предоставили несколько фенотипов (измерения формы носа, пигментацию кожи, рост, вес и ИМТ), а также соответствующие ковариаты (компоненты происхождения, пол и возраст). Там, где это уместно, мы также предоставили климатические значения (среднегодовая температура, годовой индекс засушливости и ультрафиолетовое излучение B), приписываемые каждому человеку. Эти данные могут быть использованы для воспроизведения почти всех результатов, представленных в рукописи, без ущерба для конфиденциальности участников. Некоторые необработанные данные, такие как 3D-фотографии, генотипы и места рождения, не были переданы, потому что они легко идентифицируются по своей природе, и их передача будет этическим и юридическим нарушением информированного согласия, полученного от участников.Это ограничение не связано с какими-либо личными или коммерческими интересами.
Там, где это уместно, мы также предоставили климатические значения (среднегодовая температура, годовой индекс засушливости и ультрафиолетовое излучение B), приписываемые каждому человеку. Эти данные могут быть использованы для воспроизведения почти всех результатов, представленных в рукописи, без ущерба для конфиденциальности участников. Некоторые необработанные данные, такие как 3D-фотографии, генотипы и места рождения, не были переданы, потому что они легко идентифицируются по своей природе, и их передача будет этическим и юридическим нарушением информированного согласия, полученного от участников.Это ограничение не связано с какими-либо личными или коммерческими интересами.
Финансирование: Эта работа частично поддержана Национальным институтом юстиции США, Министерством обороны США, Исследовательским грантом Междисциплинарной инновационной инициативы Университета Иллинойса, Фламандским институтом содействия инновациям в науке и технологиях. во Фландрии (IWT Vlaanderen), Исследовательская программа Фонда научных исследований — Фландрия (Бельгия) и Центр эволюции и развития человека в Пенсильвании. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Форма носа, как и многих других частей лица, варьируется как внутри, так и среди людей. Например, расстояние между носовыми крыльями (крыльями носа) значительно больше у людей западноафриканского, южноазиатского и восточноазиатского происхождения по сравнению с людьми европейского происхождения [1].Носовой указатель (ширина / высота носового отверстия черепа) также, как известно, значительно различается в разных популяциях [1,2]. Неясно, вызваны ли эти популяционные различия в форме носа в первую очередь генетическим дрейфом или естественным отбором.
Жизненно важная функция носа — нагревать вдыхаемый воздух до температуры тела и насыщать его водяным паром, прежде чем он достигнет нижних дыхательных путей [3]. Фактически, вдыхаемый воздух достигает 90% от требуемых уровней температуры и влажности еще до того, как достигает носоглотки, что делает полость носа главным кондиционирующим аппаратом в дыхательных путях [4,5].Это кондиционирование поддерживает надлежащее функционирование мукоцилиарного аппарата, который улавливает частицы и патогены и удаляет их из дыхательных путей. Низкая влажность в дыхательных путях приводит к нарушению мукоцилиарной функции и увеличивает риск инфекций как верхних, так и нижних дыхательных путей [6]. Большая часть кондиционирования воздуха происходит, когда он проходит через носовые раковины, стенки которых выстланы кровеносными сосудами и слизью, производящими бокаловидные клетки [4]. Исследования показали, что эффективность процесса кондиционирования зависит от динамики потока вдыхаемого воздуха, которая, в свою очередь, зависит от геометрии носовой полости и входных отверстий [5,7].Поскольку нос выполняет функцию кондиционера, предполагается, что различия в форме носа у разных групп населения могли быть вызваны местной адаптацией к климату [2,3,8].
Есть несколько проблем при проверке этой гипотезы. Мы знаем, что форма носа существенно различается среди человеческих популяций, как по внешней морфологии носа, так и по основной морфологии черепа [1,2,9]. Хотя это можно объяснить адаптацией к местному давлению отбора, это также можно объяснить тем фактом, что фенотипические различия между географически удаленными популяциями могут возникать просто из-за генетического дрейфа.Таким образом, чтобы использовать дивергентный отбор в качестве объяснения, необходимо продемонстрировать, что наблюдаемые вариации формы носа в разных популяциях людей больше, чем ожидаемые при одном генетическом дрейфе. Это можно сделать с помощью статистики Qst, которая измеряет степень генетической дифференциации, лежащей в основе количественного признака [10]. В принципе, ожидается, что Qst нейтрально эволюционирующего признака будет соответствовать распределению Fst нейтрально развивающихся локусов [11]. Таким образом, когда Qst намного больше, чем Fst, расхождение признаков превышает нейтральные ожидания и может быть отнесено на счет дивергентного отбора [11]. Проблема с Qst заключается в том, что его расчет требует знания аддитивных генетических вариаций внутри и между популяциями. Их можно надежно оценить только с помощью экспериментов «обычного сада», в которых влияние окружающей среды на фенотип можно эффективно контролировать. Поскольку такие эксперименты невозможны на людях, вывод на основе Qst относительно дивергентного отбора по фенотипам человека требует принятия реалистичных предположений о наследуемости рассматриваемых фенотипов.
В нескольких исследованиях использовался этот подход и было обнаружено, что хотя большинство аспектов черепа, кажется, эволюционируют нейтрально, форма носового отверстия, по-видимому, более дифференцирована в разных популяциях людей, чем ожидалось при генетическом дрейфе [9,12,13].Совсем недавно также сообщалось, что расхождение в форме внешнего носа, по крайней мере, между европейцами и китайцами-ханьцами, также превышает нейтральные ожидания [14]. Хотя это может быть правдой, в большинстве этих исследований использовались антиконсервативные предположения о наследственности, которые переоценивают генетическую дифференциацию, лежащую в основе признака, и, таким образом, приводят к неверным выводам относительно относительной роли отбора и дрейфа в движущих фенотипических различиях между популяциями.
Здесь мы провели исследование вариаций формы носа снаружи в свете количественной генетической теории, чтобы выяснить, превышает ли вариация формы носа среди популяций нейтральные ожидания.При этом мы обсуждаем методологические проблемы, связанные с решением таких вопросов у людей, и показываем, как ограничения предыдущих исследований могут быть устранены с помощью последних достижений в статистической генетике. В качестве принципиального доказательства мы сравниваем дифференциацию формы носа с двумя сильно наследуемыми морфологическими признаками, которые, как известно, имеют полигенную основу; а именно рост и пигментация кожи [15–18]. Как рост, так и пигментация кожи существенно различаются внутри и между популяциями людей, и, как полагают, в различных популяциях они подвергаются положительному отбору [19–22].Пигментация кожи также является хорошим примером фенотипа, который, как известно, развился в ответ на меняющееся в географическом пространстве давление отбора: воздействие ультрафиолетового излучения B (UVB) на уровне земли [23]. Наконец, мы проверяем, зависит ли клинальная вариация в аспектах формы носа, которая, по-видимому, ускоряется в разных популяциях, от географических вариаций температуры и влажности, чтобы определить, вызвано ли это расхождение давлением климатического отбора.
Наконец, мы проверяем, зависит ли клинальная вариация в аспектах формы носа, которая, по-видимому, ускоряется в разных популяциях, от географических вариаций температуры и влажности, чтобы определить, вызвано ли это расхождение давлением климатического отбора.
Результаты
Описание изменения формы носа
Чтобы количественно оценить вариации формы носа, мы сначала сделали трехмерные изображения лиц участников с высоким разрешением, используя систему 3dMD Face ( 3dMD Atlanta , GA ).Были размещены пять ориентиров (два на внутреннем углу глаз, два на внешних углах рта и один на кончике носа), чтобы установить ориентацию лица. Пространственно плотная сетка из 7 150 квази-ориентиров (QL) была нанесена на каждое изображение и его отражение. Обобщенное наложение Прокруста [24] было выполнено на обоих наборах изображений (исходном и отраженном) для устранения различий в положении и ориентации. Координаты Прокруста исходного и отраженного изображений для каждого участника затем были усреднены, чтобы удалить эффекты двусторонней асимметрии, как указано в Claes et al . (2014) [25]. Область носа, которая состоит из 709 из 7 150 QL, была выбрана для последующих анализов.
(2014) [25]. Область носа, которая состоит из 709 из 7 150 QL, была выбрана для последующих анализов.
Мы использовали линейные расстояния и площади, чтобы охарактеризовать форму носа (рис. 1). Линейные расстояния (измеренные в мм) были рассчитаны с использованием семи стандартных антропометрических ориентиров: i. назион (п), II. пронасале (прн), iii. subnasale (sn), iv. Кривизна левой крыльчатки (al -1 ), v. Искривление правой крыльчатки (al r ), vi. левое основание сигнализатора (ac l ) и vii.Правое основание крыльев (ac r ) (рис. 1) [26,27]. Эти семь ориентиров были размещены на пространственно-плотной сетке из 709 QL, что облегчило автоматическое размещение на каждой грани. Мы использовали координаты x, y и z этих ориентиров для вычисления пяти евклидовых расстояний: i. ширина ноздрей (al l −al r ), ii. ширина основания alar (ac l −ac r ), iii. высота носа (n – sn), iv. длина носового гребня (n – prn) и v.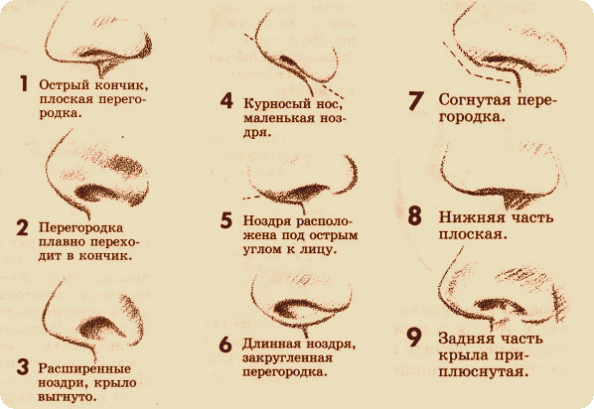 выступ кончика носа (sn – prn) (Рис. 1). Мы также вычислили две площади (измеренные в мм 2 ): i.общая внешняя площадь носа и ii. средняя площадь ноздрей (рис. 1) (методы).
выступ кончика носа (sn – prn) (Рис. 1). Мы также вычислили две площади (измеренные в мм 2 ): i.общая внешняя площадь носа и ii. средняя площадь ноздрей (рис. 1) (методы).
Рис. 1. Измерения, используемые для описания формы носа.
A) Места ориентиров, которые использовались для расчета линейных расстояний, показаны красными точками. Б) Для описания формы носа использовались пять линейных расстояний (красные линии) и две площади поверхности (красная сетка). Линейные расстояния рассчитывались по трехмерным координатам ориентиров (красные точки). Площадь поверхности рассчитывалась как сумма многоугольников, выделенных красным.
https://doi.org/10.1371/journal.pgen.1006616.g001
На рис. 2 показано распределение аспектов формы носа у мужчин и женщин из четырех групп населения: Западной Африки, Восточной Азии, Южной Азии и Северной Европейский. Эти группы были определены на основе генетического и самооценочного происхождения (методы). На рис. 2 для сравнения также показано распределение роста и индекса меланина, показателя уровня пигментации кожи, полученного на основе спектрофотометрии отражательной способности (измеренной на внутренней стороне верхней части рук; см. «Методы»).Размер выборки, среднее значение и стандартное отклонение для каждого фенотипа представлены в таблице 1. Одно четкое наблюдение состоит в том, что все аспекты формы носа, по крайней мере, рассматриваемые здесь, в значительной степени сексуально диморфны (рис. 2, таблица 1). У мужчин в среднем более широкие ноздри, более длинные носовые гребни, более выступающие наружу кончики носа, более крупные ноздри и большая общая площадь внешней поверхности по сравнению с женщинами (рис. 2, таблица 1). Все размеры носа также значительно различаются в разных популяциях (таблица S1).На основании таблицы 1 отметим некоторые общие закономерности. Например, ширина ноздрей и ширина крылового основания самые большие у западноафриканцев и самые маленькие у северных европейцев.
На рис. 2 для сравнения также показано распределение роста и индекса меланина, показателя уровня пигментации кожи, полученного на основе спектрофотометрии отражательной способности (измеренной на внутренней стороне верхней части рук; см. «Методы»).Размер выборки, среднее значение и стандартное отклонение для каждого фенотипа представлены в таблице 1. Одно четкое наблюдение состоит в том, что все аспекты формы носа, по крайней мере, рассматриваемые здесь, в значительной степени сексуально диморфны (рис. 2, таблица 1). У мужчин в среднем более широкие ноздри, более длинные носовые гребни, более выступающие наружу кончики носа, более крупные ноздри и большая общая площадь внешней поверхности по сравнению с женщинами (рис. 2, таблица 1). Все размеры носа также значительно различаются в разных популяциях (таблица S1).На основании таблицы 1 отметим некоторые общие закономерности. Например, ширина ноздрей и ширина крылового основания самые большие у западноафриканцев и самые маленькие у северных европейцев. Эти результаты согласуются с предыдущими наблюдениями [1,2]. Выступ кончика носа больше всего у североевропейцев и меньше всего у западноафриканцев и восточноазиатцев. E. Азиаты также имеют самые маленькие носы с точки зрения площади внешней поверхности.
Эти результаты согласуются с предыдущими наблюдениями [1,2]. Выступ кончика носа больше всего у североевропейцев и меньше всего у западноафриканцев и восточноазиатцев. E. Азиаты также имеют самые маленькие носы с точки зрения площади внешней поверхности.
Рис. 2. Коробчатые диаграммы фенотипов по популяции и полу, наложенные на отдельные точки данных.
Рост измеряется в сантиметрах, а индекс меланина измеряется в процентах отражательной способности (методы).Линейные расстояния измеряются в миллиметрах (мм), а площадь измеряется в мм. 2 . Точки представляют собой индивидуальные наблюдения, а цвет прямоугольных диаграмм и точек представляет пол, причем синий цвет указывает на мужчин, а красный — на женщин.
https://doi.org/10.1371/journal.pgen.1006616.g002
Тестирование на ускоренную дивергенцию
Различия в фенотипах могут накапливаться в разных популяциях просто из-за генетического дрейфа. Чтобы вызвать положительный направленный отбор, нужно продемонстрировать, что вариации между популяциями больше, чем ожидаемые при генетическом дрейфе. Мы использовали Qst (см. Недавний обзор Leinonen и др. [28]), количественный генетический аналог Fst [10,29], чтобы проверить, проявляют ли определенные аспекты формы носа большую дифференциацию между популяциями, чем ожидалось при генетическом дрейфе. в одиночестве. Qst измеряет степень генетической дифференциации количественного признака в популяциях и определяется как:
(1)
где и — компоненты фенотипической дисперсии из-за аддитивных генетических эффектов среди и внутри популяций, соответственно.Было показано, что, в принципе, ожидается, что распределение Qst количественного признака, которое развилось только в результате генетического дрейфа, будет равно Fst нейтральных генетических маркеров [11,30,31]. Это ожидание позволяет сравнивать Qst с Fst, чтобы проверить, достаточно ли одного генетического дрейфа для объяснения дивергенции признака среди популяций. Если Qst признака в наборе популяций намного больше, чем Fst, это означает, что фенотипическая дифференциация превышает ожидание при нейтральности.
Мы использовали Qst (см. Недавний обзор Leinonen и др. [28]), количественный генетический аналог Fst [10,29], чтобы проверить, проявляют ли определенные аспекты формы носа большую дифференциацию между популяциями, чем ожидалось при генетическом дрейфе. в одиночестве. Qst измеряет степень генетической дифференциации количественного признака в популяциях и определяется как:
(1)
где и — компоненты фенотипической дисперсии из-за аддитивных генетических эффектов среди и внутри популяций, соответственно.Было показано, что, в принципе, ожидается, что распределение Qst количественного признака, которое развилось только в результате генетического дрейфа, будет равно Fst нейтральных генетических маркеров [11,30,31]. Это ожидание позволяет сравнивать Qst с Fst, чтобы проверить, достаточно ли одного генетического дрейфа для объяснения дивергенции признака среди популяций. Если Qst признака в наборе популяций намного больше, чем Fst, это означает, что фенотипическая дифференциация превышает ожидание при нейтральности.
Компоненты аддитивной генетической изменчивости и, как правило, оцениваются на основе экспериментов «общего сада», в которых можно контролировать влияние окружающей среды [28]. Это часто невозможно для немодельных организмов, особенно для людей, из-за практических и этических ограничений. В таких случаях и может быть оценено на основе внутрипопуляционных и внутрипопуляционных компонентов фенотипической дисперсии и, соответственно, при условии, что наследуемость, лежащая в основе этих компонентов, известна: (2)
Здесь и — внутрипопуляционные компоненты фенотипической дисперсии, а c и h 2 — пропорции и, соответственно, обусловленные аддитивными генетическими эффектами.И c , и h 2 могут варьироваться от 0 (ни одна из вариаций не обусловлена аддитивными генетическими эффектами) до 1 (вся вариация обусловлена аддитивными генетическими эффектами). Уравнение (2) показывает, что Qst, вычисленное из компонентов фенотипической дисперсии, зависит от соотношения между c и h 2 [32]. Без предварительной информации разумно предположить, что ц / ч 2 = 1, т.е. доля фенотипической дисперсии, обусловленная аддитивными генетическими эффектами, одинакова среди популяций и внутри популяций.Рассчитанный таким образом Qst иногда называют Pst [33]. Однако мы продолжим использовать термин Qst, чтобы избежать путаницы, и оценим справедливость предположения, что c / h 2 = 1 в следующем разделе.
Без предварительной информации разумно предположить, что ц / ч 2 = 1, т.е. доля фенотипической дисперсии, обусловленная аддитивными генетическими эффектами, одинакова среди популяций и внутри популяций.Рассчитанный таким образом Qst иногда называют Pst [33]. Однако мы продолжим использовать термин Qst, чтобы избежать путаницы, и оценим справедливость предположения, что c / h 2 = 1 в следующем разделе.
Мы рассчитали Qst для каждого аспекта формы носа, описанного в предыдущем разделе, по четырем группам населения: i) западноафриканский (N = 40), ii) североевропейский (N = 236), iii) восточноазиатский (N = 127) и iv) Южная Азия (N = 73) (см. Методы критериев отбора).Мы использовали непараметрический подход начальной загрузки для создания эмпирических распределений Qst и Fst и для проверки того, превышает ли наблюдаемое значение Qst значение Fst (методы). Мы использовали статистику Qst – Fst, которая, согласно нулевой гипотезе генетического дрейфа, должна быть равна нулю. Чем больше Qst – Fst фенотипа, тем сильнее доказательство того, что вариации фенотипа в популяциях больше, чем ожидаемые при одном только генетическом дрейфе. Мы называем выбросы в нейтральном распределении сигналами ускоренной дивергенции для краткости.Сила доказательства ускоренной дивергенции может быть измерена с помощью эмпирического p-значения, которое представляет собой долю бутстрепных значений Qst – Fst, которые меньше нуля. Чтобы сравнить с другими количественными признаками на полигенной основе, мы также проверили, являются ли рост и пигментация кожи сигналами ускоренной дивергенции. Результаты показаны на рис. 3, а значения p перечислены в таблице 2. Мы обрабатываем фенотипы, прошедшие строгую поправку Бонферонни (значение p <0,0071 = 0.05/7 для семи черт формы носа) как свидетельство ускоренного расхождения между популяциями.
Чем больше Qst – Fst фенотипа, тем сильнее доказательство того, что вариации фенотипа в популяциях больше, чем ожидаемые при одном только генетическом дрейфе. Мы называем выбросы в нейтральном распределении сигналами ускоренной дивергенции для краткости.Сила доказательства ускоренной дивергенции может быть измерена с помощью эмпирического p-значения, которое представляет собой долю бутстрепных значений Qst – Fst, которые меньше нуля. Чтобы сравнить с другими количественными признаками на полигенной основе, мы также проверили, являются ли рост и пигментация кожи сигналами ускоренной дивергенции. Результаты показаны на рис. 3, а значения p перечислены в таблице 2. Мы обрабатываем фенотипы, прошедшие строгую поправку Бонферонни (значение p <0,0071 = 0.05/7 для семи черт формы носа) как свидетельство ускоренного расхождения между популяциями.
Рис. 3. Результаты Qst – Fst для всех популяций.
Бутстрепное распределение Qst – Fst для каждого фенотипа (показанное графиком скрипки) сравнивается с ожидаемым нулевым значением при нейтральности (горизонтальная пунктирная линия). Фенотипы, которые демонстрируют ускоренную дивергенцию (с использованием порогового значения p, скорректированного по Бонферронни, равного 0,0071), показаны красным.
Фенотипы, которые демонстрируют ускоренную дивергенцию (с использованием порогового значения p, скорректированного по Бонферронни, равного 0,0071), показаны красным.
https: // doi.org / 10.1371 / journal.pgen.1006616.g003
Как и ожидалось, пигментация кожи свидетельствует об ускорении дивергенции между популяциями (Qst = 0,642, значение p = 1,00E-04). Это соответствует идее о том, что различия в пигментации кожи, по крайней мере, среди континентальных популяций, были вызваны положительным отбором. Высота, с другой стороны, не показывает сигналов об ускоренной дивергенции (Qst = 0,177, p-value = 0,14). Хотя рост, по-видимому, находится под положительным отбором в пределах европейских и африканских популяций отдельно [19,34], он, по-видимому, не вызван дивергентным давлением отбора в рассматриваемых здесь популяциях.Из всех измерений формы носа только ширина ноздри (Qst = 0,467, значение p = 2,80E-03) и ширина основания носа (Qst = 0,440, значение p = 4,90E-03) показывают сигналы ускоренной дивергенции. Однако степень расхождения этих признаков не так высока, как наблюдаемая для пигментации кожи.
Однако степень расхождения этих признаков не так высока, как наблюдаемая для пигментации кожи.
Оценка обоснованности предположений относительно
c и h 2 Мы оценили Qst в предположении, что ц / ч 2 = 1.Другими словами, мы предполагаем, что доля межпопуляционной дисперсии признака из-за аддитивных генетических эффектов такая же, как доля внутрипопуляционной дисперсии из-за аддитивных генетических эффектов. Это может быть неверным предположением, если большая часть фенотипической дисперсии между популяциями обусловлена прямым воздействием окружающей среды (т.е. фенотипической пластичностью). Если истинное значение ц / ч 2 значительно ниже 1, полученные по фенотипу оценки генетической дисперсии между популяциями и, следовательно, Qst будут завышены, что приведет к ложноположительным выводам относительно роли отбора в управлении фенотипической дифференциацией. Этот факт в значительной степени игнорировался в предыдущих исследованиях, в которых подходы на основе Qst использовались для изучения краниофациальной дивергенции [12–14,35]. В этих исследованиях h 2 было принято равным 0,55, а c неявно предполагалось равным 1, в результате чего c / h 2 было больше 1. Хотя это могло быть верно для некоторых признаков (например, см. пигментацию кожи в Таблице 3), это, по нашему мнению, антиконсервативный подход, поскольку он предполагает, что наследуемость между популяциями выше, чем наследуемость внутри популяций.Во всяком случае, противоположный случай ( c / h 2 <1) более правдоподобен, поскольку мы ожидаем, что экологические вариации будут большими среди географически удаленных популяций. Имея это в виду, мы оценили чувствительность результатов Qst – Fst к случаю, когда c / h 2 <1 [32], определив «критическое значение» c / h 2 , при котором 95% нижняя граница Qst соответствует 95% верхней границе Fst (рис.
Этот факт в значительной степени игнорировался в предыдущих исследованиях, в которых подходы на основе Qst использовались для изучения краниофациальной дивергенции [12–14,35]. В этих исследованиях h 2 было принято равным 0,55, а c неявно предполагалось равным 1, в результате чего c / h 2 было больше 1. Хотя это могло быть верно для некоторых признаков (например, см. пигментацию кожи в Таблице 3), это, по нашему мнению, антиконсервативный подход, поскольку он предполагает, что наследуемость между популяциями выше, чем наследуемость внутри популяций.Во всяком случае, противоположный случай ( c / h 2 <1) более правдоподобен, поскольку мы ожидаем, что экологические вариации будут большими среди географически удаленных популяций. Имея это в виду, мы оценили чувствительность результатов Qst – Fst к случаю, когда c / h 2 <1 [32], определив «критическое значение» c / h 2 , при котором 95% нижняя граница Qst соответствует 95% верхней границе Fst (рис. 4).Меньшие критические значения означают, что сравнения Qst – Fst на основе фенотипа устойчивы к предположениям ц / ч 2 . Такие «анализы чувствительности», хотя и распространены в литературе по молекулярной экологии, недостаточно представлены в генетических и антропологических исследованиях человека. На рис. 4 показаны кривые чувствительности к признакам роста, пигментации кожи и формы носа. Пигментация кожи, которая показывает очень высокое значение Qst, имеет критическое значение 0,25, что намного ниже 1 (рис. 4 и таблица 2).Критическое значение ширины ноздрей составляет 0,50, а ширина крылового основания — 0,55, что также намного меньше 1 (Рис. 4 и Таблица 2). Таким образом, мы ожидаем, что эти фенотипы будут демонстрировать сигналы ускоренной дивергенции, даже если истинные значения ц / ч 2 были намного меньше, чем 1.
4).Меньшие критические значения означают, что сравнения Qst – Fst на основе фенотипа устойчивы к предположениям ц / ч 2 . Такие «анализы чувствительности», хотя и распространены в литературе по молекулярной экологии, недостаточно представлены в генетических и антропологических исследованиях человека. На рис. 4 показаны кривые чувствительности к признакам роста, пигментации кожи и формы носа. Пигментация кожи, которая показывает очень высокое значение Qst, имеет критическое значение 0,25, что намного ниже 1 (рис. 4 и таблица 2).Критическое значение ширины ноздрей составляет 0,50, а ширина крылового основания — 0,55, что также намного меньше 1 (Рис. 4 и Таблица 2). Таким образом, мы ожидаем, что эти фенотипы будут демонстрировать сигналы ускоренной дивергенции, даже если истинные значения ц / ч 2 были намного меньше, чем 1.
Рис. 4. Графики чувствительности, показывающие критические значения c / h 2 .
Изменение медианы Qst роста, пигментации кожи и формы носа в зависимости от c / h 2 показано сплошной красной линией.Медиана Fst показана сплошной синей линией. Нижняя и верхняя 95% границы для Qst и Fst показаны пунктирными красными и синими линиями соответственно. Критическое значение, при котором нижняя граница Qst встречается с верхней границей Fst, показано в верхнем левом углу каждого графика. Более низкие критические значения указывают на то, что Qst – Fst более устойчив к предположениям c / h 2 .
https://doi.org/10.1371/journal.pgen.1006616.g004
Изучение наследственности формы носа внутри и среди популяций
В дополнение к анализу чувствительности мы хотели продемонстрировать, что различия в форме носа у людей как внутри, так и между популяциями являются наследственными.Внутрипопуляционная наследственность ( h 2 ) по фенотипу традиционно оценивается на основе данных, собранных на больших группах близнецов или родословных, в которых известны генетические отношения между особями [36]. Недавно Янг и его коллеги (2010) представили линейный подход смешанной модели, который можно использовать для оценки альтернативной статистики у неродственных людей: доля фенотипической дисперсии, объясняемая генотипированными SNP ( h g 2 ) [18].Основное преимущество этого подхода перед традиционными подходами, основанными на близнецах и родословных, заключается в том, что он применим к данным от неродственных лиц, которые более доступны и легче собирать в больших количествах. Мы вычислили h g 2 как оценку h 2 для роста, индекса меланина и формы носа, используя 118 420 аутосомных SNP в выборке неродственных людей европейского происхождения. , реализованный в программе GCTA [37] (Методы).Значения h g 2 представлены в таблице 3. Наследуемость высоты у европейцев ( h g 2 = 0,394; N = 1,825) аналогичны ранее опубликованным оценкам [18].
Недавно Янг и его коллеги (2010) представили линейный подход смешанной модели, который можно использовать для оценки альтернативной статистики у неродственных людей: доля фенотипической дисперсии, объясняемая генотипированными SNP ( h g 2 ) [18].Основное преимущество этого подхода перед традиционными подходами, основанными на близнецах и родословных, заключается в том, что он применим к данным от неродственных лиц, которые более доступны и легче собирать в больших количествах. Мы вычислили h g 2 как оценку h 2 для роста, индекса меланина и формы носа, используя 118 420 аутосомных SNP в выборке неродственных людей европейского происхождения. , реализованный в программе GCTA [37] (Методы).Значения h g 2 представлены в таблице 3. Наследуемость высоты у европейцев ( h g 2 = 0,394; N = 1,825) аналогичны ранее опубликованным оценкам [18]. Интересно, что наследуемость пигментации кожи у европейцев очень низкая ( h g 2 = 0,191; N = 1231), что, по нашему мнению, можно объяснить снижением генетической изменчивости локусов пигментации кожи из-за сильный положительный отбор для более светлой кожи у европейцев [17,21,38].Почти все аспекты формы носа, по-видимому, в значительной степени наследуются у европейцев ( h g 2 диапазон: 0,401–0,657 N = 1,718) (Таблица 3).
Интересно, что наследуемость пигментации кожи у европейцев очень низкая ( h g 2 = 0,191; N = 1231), что, по нашему мнению, можно объяснить снижением генетической изменчивости локусов пигментации кожи из-за сильный положительный отбор для более светлой кожи у европейцев [17,21,38].Почти все аспекты формы носа, по-видимому, в значительной степени наследуются у европейцев ( h g 2 диапазон: 0,401–0,657 N = 1,718) (Таблица 3).
Оценка наследуемости среди популяций ( c ) более трудна, поскольку генетические эффекты между географически удаленными популяциями могут быть смешаны с большими экологическими эффектами. Этот параметр часто можно надежно оценить только с помощью экспериментов «общего сада», в которых могут быть приняты во внимание систематические различия в окружающей среде, которые могут быть смешаны с генетическими различиями.Однако, как упоминалось ранее, этот подход возможен только в модельных организмах, которые поддаются экспериментальным манипуляциям. У людей и других немодельных организмов это серьезное ограничение, которое можно преодолеть, изучая встречающиеся в природе смешанные популяции.
У людей и других немодельных организмов это серьезное ограничение, которое можно преодолеть, изучая встречающиеся в природе смешанные популяции.
Процесс смешения объединяет генофонды двух или более популяций, которые могли разойтись из-за генетического дрейфа, мутации и отбора. Смешивание, которое может происходить неоднократно в течение нескольких поколений, с последующей рекомбинацией, приводит к хромосомам, которые представляют собой мозаику сегментов предков (Рис. 5) [39].В случайно спаривающейся смешанной популяции сегменты родословной разделяются случайным образом по отношению к окружающей среде, что отделяет межпопуляционные генетические эффекты на фенотип от эффектов окружающей среды, что позволяет оценить генетическую вариативность, лежащую в основе фенотипа. Мы предполагаем, что метод, недавно разработанный для оценки наследуемости в смешанных популяциях (Zaitlen et al . (2014)) [40], может дать достоверную оценку c . Зайтлен и др. .(2014) расширяют метод, разработанный Янгом и др. . (2010) [18], используя локальную родословную в SNP вместо генотипов, чтобы построить матрицу генетических отношений между смешанными особями. Доля фенотипической дисперсии в смешанной популяции, которая может быть объяснена местным происхождением ( h y 2 ), концептуально эквивалентна c между родительскими популяциями. Мы рассуждаем о том, что на генотипическом уровне генетическая изменчивость в смешанной популяции должна быть суммой генетической изменчивости в родительских популяциях и генетической изменчивости между ними.Вариации в масштабе местной родословной представляют только генетические различия между популяциями (рис. 5). Таким образом, доля фенотипической изменчивости, которая может быть объяснена местным происхождением ( h y 2 ), должна быть эквивалентна c .
. (2010) [18], используя локальную родословную в SNP вместо генотипов, чтобы построить матрицу генетических отношений между смешанными особями. Доля фенотипической дисперсии в смешанной популяции, которая может быть объяснена местным происхождением ( h y 2 ), концептуально эквивалентна c между родительскими популяциями. Мы рассуждаем о том, что на генотипическом уровне генетическая изменчивость в смешанной популяции должна быть суммой генетической изменчивости в родительских популяциях и генетической изменчивости между ними.Вариации в масштабе местной родословной представляют только генетические различия между популяциями (рис. 5). Таким образом, доля фенотипической изменчивости, которая может быть объяснена местным происхождением ( h y 2 ), должна быть эквивалентна c .
Рис. 5. Смесь объединяет внутрипопуляционные и межпопуляционные различия у смешанных особей.
Генетическая изменчивость между двумя родительскими популяциями представлена различием в цвете хромосом, тогда как генетическая изменчивость внутри популяций представлена интенсивностью оттенка.Смесь объединяет генетические вариации обеих популяций. На генотипическом уровне генетическая изменчивость в смешанных популяциях состоит как из внутрипопуляционных, так и из межпопуляционных вариаций. Различия на уровне предков — это вариации между двумя родительскими популяциями.
https://doi.org/10.1371/journal.pgen.1006616.g005
Следуя этим рассуждениям, мы оценили h y 2 от местной родословной на 623 625 аутосомных SNP в выборке из 409 Жители Кабо-Верде, которые произошли в основном от W.Африканцы и европейцы (см. «Методы» для получения более подробной информации и Таблицу 3 для результатов). Таблица 3 показывает, что фенотипические различия между западными африканцами и европейцами по росту ( h y 2 = 0,224, N = 409) вполне наследуемы. Результаты для данных о пигментации кожи здесь не представлены, так как они были недоступны. Различия во многих аспектах формы носа также наследуются (ширина ноздри: h y 2 = 0.226, Ширина основания носа: h y 2 = 0,212, Выступ кончика носа: 0,177, Площадь внешней поверхности: h y 2 = 0,121, N = 409). Другие аспекты формы носа могут не передаваться по наследству между этими двумя популяциями (высота носа: h y 2 = 0,03, h y 2 = 0.059, N = 409). Еще одно интересное наблюдение из таблицы 3 состоит в том, что оценки h y 2 обычно ниже, чем оценки h g 2 для всех признаков. Это предполагает, что для большинства человеческих черт аддитивная генетическая дисперсия между популяциями может быть меньше аддитивной генетической дисперсии внутри популяций, что согласуется с тем фактом, что большая часть генетической изменчивости у людей существует внутри популяций.
Результаты для данных о пигментации кожи здесь не представлены, так как они были недоступны. Различия во многих аспектах формы носа также наследуются (ширина ноздри: h y 2 = 0.226, Ширина основания носа: h y 2 = 0,212, Выступ кончика носа: 0,177, Площадь внешней поверхности: h y 2 = 0,121, N = 409). Другие аспекты формы носа могут не передаваться по наследству между этими двумя популяциями (высота носа: h y 2 = 0,03, h y 2 = 0.059, N = 409). Еще одно интересное наблюдение из таблицы 3 состоит в том, что оценки h y 2 обычно ниже, чем оценки h g 2 для всех признаков. Это предполагает, что для большинства человеческих черт аддитивная генетическая дисперсия между популяциями может быть меньше аддитивной генетической дисперсии внутри популяций, что согласуется с тем фактом, что большая часть генетической изменчивости у людей существует внутри популяций. Тем не менее, мы осторожно переоцениваем этот вывод, поскольку наследственность была оценена только у людей с западноафриканским и европейским происхождением и не отражала вариации внутри и между другими популяциями. В целом наши результаты показывают, что генетические различия лежат в основе различий во многих аспектах формы носа как внутри, так и между популяциями.
Тем не менее, мы осторожно переоцениваем этот вывод, поскольку наследственность была оценена только у людей с западноафриканским и европейским происхождением и не отражала вариации внутри и между другими популяциями. В целом наши результаты показывают, что генетические различия лежат в основе различий во многих аспектах формы носа как внутри, так и между популяциями.
Тестирование на адаптацию к климату
Ранее несколько исследований показали, что форма носовой апертуры и носовой полости коррелирует с климатическими переменными, связанными с температурой и влажностью, так что у людей из холодно-сухого климата носовые полости более узкие по сравнению с людьми из теплого влажного климата [41, 42].Мы были заинтересованы в том, чтобы проверить, демонстрируют ли аспекты формы внешнего носа, демонстрирующие необычно высокую дифференциацию среди популяций, на основе анализа Qst – Fst, корреляции с климатом. Для этой цели мы выбрали из подмножества, используемого в анализе Qst, женщин с генетическими данными, родители которых родились в регионе, который совпадает с их континентальной родословной (N = 140) (Рис. 6). Это было сделано для того, чтобы присвоить каждому человеку климатическую ценность, наиболее близкую к климату их «предков». Поскольку у нас не было данных о генотипах мужчин североевропейского происхождения, мы использовали только женщин для этого анализа.Данные генотипа были необходимы для корректировки генетической структуры (см. Методы). Мы проверили корреляцию ширины ноздрей и ширины крылового основания с тремя климатическими переменными: i) среднегодовой температурой (далее именуемой температурой), ii) относительной влажностью и iii) абсолютной влажностью. Выбор этих климатических переменных следует из функционального значения носа в нагревании и увлажнении вдыхаемого воздуха. Мы также проверили, связана ли пигментация кожи с уровнем УФ-В. Это было использовано в качестве доказательства принципа, поскольку многочисленные свидетельства предполагают, что различия в пигментации кожи в разных популяциях развивались в первую очередь в ответ на ультрафиолетовое излучение [17, 20–22].
6). Это было сделано для того, чтобы присвоить каждому человеку климатическую ценность, наиболее близкую к климату их «предков». Поскольку у нас не было данных о генотипах мужчин североевропейского происхождения, мы использовали только женщин для этого анализа.Данные генотипа были необходимы для корректировки генетической структуры (см. Методы). Мы проверили корреляцию ширины ноздрей и ширины крылового основания с тремя климатическими переменными: i) среднегодовой температурой (далее именуемой температурой), ii) относительной влажностью и iii) абсолютной влажностью. Выбор этих климатических переменных следует из функционального значения носа в нагревании и увлажнении вдыхаемого воздуха. Мы также проверили, связана ли пигментация кожи с уровнем УФ-В. Это было использовано в качестве доказательства принципа, поскольку многочисленные свидетельства предполагают, что различия в пигментации кожи в разных популяциях развивались в первую очередь в ответ на ультрафиолетовое излучение [17, 20–22]. Связь между фенотипами и климатическими переменными была проверена с использованием линейных смешанных моделей, которые корректируют возраст, ИМТ и генетическое сходство (методы). Пол не был включен в качестве ковариаты, поскольку использовались только самки. Мы использовали тест отношения правдоподобия (LRT), чтобы оценить статистическую значимость наклона между фенотипом и климатической переменной. Статистика LRT и соответствующие ей p-значения были получены путем сравнения полной (включая предсказатель климата) и сокращенной (предсказатель климата удален) моделей.Результаты представлены в таблице 4.
Связь между фенотипами и климатическими переменными была проверена с использованием линейных смешанных моделей, которые корректируют возраст, ИМТ и генетическое сходство (методы). Пол не был включен в качестве ковариаты, поскольку использовались только самки. Мы использовали тест отношения правдоподобия (LRT), чтобы оценить статистическую значимость наклона между фенотипом и климатической переменной. Статистика LRT и соответствующие ей p-значения были получены путем сравнения полной (включая предсказатель климата) и сокращенной (предсказатель климата удален) моделей.Результаты представлены в таблице 4.
Рис. 6. Географическое распределение мест рождения родителей.
Отдельные точки обозначают места рождения родителей с линией, соединяющей двух родителей. Единственная точка означает, что оба родителя родились в одном месте. Значения климата в этих местах использовались для проверки сигналов об адаптации к климату.
https://doi.org/10.1371/journal.pgen. 1006616.g006
1006616.g006
Как и ожидалось, мы видим сильную взаимосвязь между пигментацией кожи и УФ-В (N = 126, T = 6.23, LRT = 55.20, p-значение = 1.09E-13). Положительные значения наклона и t-статистики указывают на то, что люди, как правило, темнее в регионах с более высоким воздействием УФ-В и наоборот. Эта закономерность дополнительно подтверждает идею о том, что адаптация к ультрафиолетовому излучению сыграла важную роль в эволюции пигментации кожи у людей. Ширина ноздрей значительно коррелирует с температурой (N = 140, T = 3,09, LRT = 9,24, p-value = 2,37E-03) и абсолютной влажностью (N = 140, T = 2,73, LRT = 7.28, p-значение = 6.97E-03). Положительные наклоны указывают на то, что люди из теплого влажного климата в среднем обычно имеют более широкую ноздрю, тогда как люди из прохладно-сухого климата, как правило, имеют более узкую ноздрю. Это согласуется с ранее описанными паттернами экогеографических вариаций ширины носовой апертуры и носовой полости [41,42]. Ширина ноздрей не имела значимой корреляции с относительной влажностью (N = 140, T = -1,65, LRT = 2,71, значение p = 0,100). Это неудивительно, поскольку абсолютная влажность более важна для физиологического функционирования носа, чем относительная влажность [43].Ширина основания крыльев существенно не коррелирует ни с температурой (N = 140, T = 1,97, LRT = 3,82, значение p = 0,051), ни с относительной влажностью (N = 140, T = -0,79, LRT = 0,62, значение p = 0,431) и лишь слегка коррелирует с абсолютной влажностью (N = 140, T = 2,09, LRT = 4,30, p-значение = 0,038). Это может означать, что не ширина носа как таковая , а ширина ноздрей находится под давлением отбора. Однако даже ширина ноздрей не так сильно коррелирует с температурой и влажностью, как пигментация кожи — с УФ-В, что позволяет предположить, что сила отбора, оказываемого температурой на ширину носа, вероятно, будет намного слабее, чем давление отбора, оказываемое воздействием УФ-В на пигментацию кожи. .
Это неудивительно, поскольку абсолютная влажность более важна для физиологического функционирования носа, чем относительная влажность [43].Ширина основания крыльев существенно не коррелирует ни с температурой (N = 140, T = 1,97, LRT = 3,82, значение p = 0,051), ни с относительной влажностью (N = 140, T = -0,79, LRT = 0,62, значение p = 0,431) и лишь слегка коррелирует с абсолютной влажностью (N = 140, T = 2,09, LRT = 4,30, p-значение = 0,038). Это может означать, что не ширина носа как таковая , а ширина ноздрей находится под давлением отбора. Однако даже ширина ноздрей не так сильно коррелирует с температурой и влажностью, как пигментация кожи — с УФ-В, что позволяет предположить, что сила отбора, оказываемого температурой на ширину носа, вероятно, будет намного слабее, чем давление отбора, оказываемое воздействием УФ-В на пигментацию кожи. .
Следуя предложению одного из составителей обзора, мы исследовали, были ли наблюдаемые корреляции фенотип-климат обусловлены какой-либо одной популяцией. Для этого мы повторно оценили наклоны фенотипа-климата (пигментация кожи в зависимости от УФ-В и ширины ноздрей в зависимости от температуры) после удаления каждой популяции по очереди. На рис. 7 показано, что 95% доверительные интервалы наклона перекрываются с нулем (красная линия на рис. 7) только при удалении N.Europeans, что позволяет предположить, что обе корреляции фенотип-климат обусловлены в первую очередь включением N.Европейцы. Этот эффект может указывать на положительный отбор для более светлой пигментации кожи и более узких носовых ходов в популяциях более высоких широт. Однако, поскольку наша выборка популяций на более высоких широтах ограничена (рис. 6), этот результат требует дальнейшего изучения.
Для этого мы повторно оценили наклоны фенотипа-климата (пигментация кожи в зависимости от УФ-В и ширины ноздрей в зависимости от температуры) после удаления каждой популяции по очереди. На рис. 7 показано, что 95% доверительные интервалы наклона перекрываются с нулем (красная линия на рис. 7) только при удалении N.Europeans, что позволяет предположить, что обе корреляции фенотип-климат обусловлены в первую очередь включением N.Европейцы. Этот эффект может указывать на положительный отбор для более светлой пигментации кожи и более узких носовых ходов в популяциях более высоких широт. Однако, поскольку наша выборка популяций на более высоких широтах ограничена (рис. 6), этот результат требует дальнейшего изучения.
Рис. 7. Влияние удаления каждой популяции на фенотип-климатические корреляции.
Точечная оценка и 95% доверительные интервалы уклонов фенотип-климат после удаления каждой популяции (по оси Y) показаны для A) пигментации кожи и УФ-излучения B, B) ширины и температуры ноздрей и C) ширины ноздрей.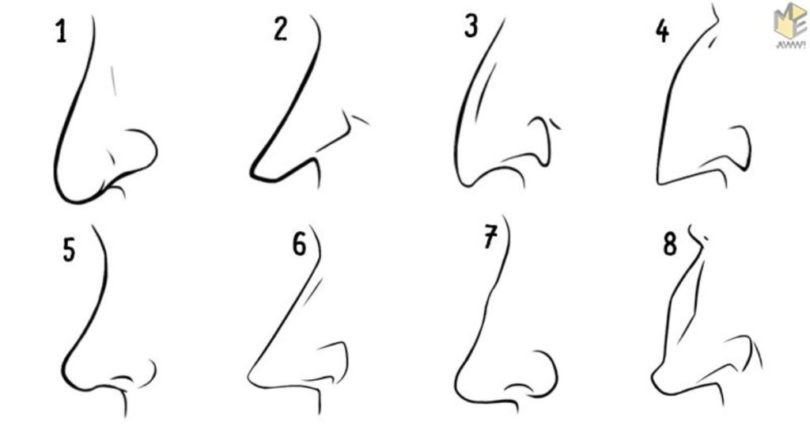 и абсолютная влажность.Во всех случаях 95% доверительный интервал наклона перекрывается с нулем (красная вертикальная линия), только когда европейцы удалены.
и абсолютная влажность.Во всех случаях 95% доверительный интервал наклона перекрывается с нулем (красная вертикальная линия), только когда европейцы удалены.
https://doi.org/10.1371/journal.pgen.1006616.g007
Обсуждение
Разнообразие черт лица в разных популяциях людей давно интересовало ученых. Несмотря на то, что генетический дрейф играл преобладающую роль в эволюции человека, внешние физические черты, такие как форма лица и пигментация кожи, из-за их близости к окружающей среде, также, вероятно, находились под влиянием естественного отбора.Накоплены существенные доказательства, подтверждающие гипотезу о том, что различия в пигментации кожи в разных популяциях человека эволюционировали в основном в ответ на давление отбора, наложенное воздействием ультрафиолетового излучения [20]. Неясно, как отбор мог повлиять на форму лица — черту, которая также весьма варьируется в разных популяциях, вероятно, потому, что до сих пор ей уделялось гораздо меньше внимания. Учитывая сложность лица, мы выбрали для изучения одну особенно интересную и изменчивую часть лица; нос.Общий вопрос, лежащий в основе этого исследования, таков: сыграла ли адаптация к климату важную роль в изменении формы человеческого носа? Чтобы ответить на этот вопрос, мы сформулировали две гипотезы: i. дивергентный отбор был вовлечен в дифференциацию определенных аспектов формы носа в разных популяциях, и ii. климат является агентом отбора в тех случаях, когда может быть задействован дивергентный отбор.
Учитывая сложность лица, мы выбрали для изучения одну особенно интересную и изменчивую часть лица; нос.Общий вопрос, лежащий в основе этого исследования, таков: сыграла ли адаптация к климату важную роль в изменении формы человеческого носа? Чтобы ответить на этот вопрос, мы сформулировали две гипотезы: i. дивергентный отбор был вовлечен в дифференциацию определенных аспектов формы носа в разных популяциях, и ii. климат является агентом отбора в тех случаях, когда может быть задействован дивергентный отбор.
Чтобы проверить первую гипотезу, мы использовали сравнения Qst – Fst, чтобы выяснить, больше ли средняя разница в форме носа среди популяций, чем ожидаемая только при генетическом дрейфе.Оценка Qst основана на учете внутрипопуляционных и межпопуляционных компонентов фенотипической дисперсии, обусловленных аддитивными генетическими эффектами. Эти компоненты дисперсии идеально рассчитываются с помощью экспериментов, в которых можно контролировать влияние переменных окружающей среды [28]. Это представляет собой практическую и этическую проблему для немодельных организмов, таких как люди, которые не поддаются требуемому типу экспериментов. Чтобы обойти эти ограничения, исследования часто рассчитывают Qst непосредственно из данных фенотипа в предположении, что наследуемость внутри популяции ( h 2 ) равна наследуемости между популяциями ( c ).Рассчитанный таким образом Qst, вероятно, лучше всего считать «минимальным» Qst, предложенным Релетфордом (1994) [35]. Однако представление о том, что здесь принимается минимальное значение Qst, основывается на том, что h 2 всегда меньше c , что, как отмечалось ранее, является антиконсервативным. Фактически, генетическая изменчивость между популяциями, вероятно, будет меньше, чем генетическая изменчивость внутри популяций для большинства фенотипов. Наш подход аналогичен предыдущим исследованиям в том, что мы также оценили Qst, приняв c = h 2 .
Это представляет собой практическую и этическую проблему для немодельных организмов, таких как люди, которые не поддаются требуемому типу экспериментов. Чтобы обойти эти ограничения, исследования часто рассчитывают Qst непосредственно из данных фенотипа в предположении, что наследуемость внутри популяции ( h 2 ) равна наследуемости между популяциями ( c ).Рассчитанный таким образом Qst, вероятно, лучше всего считать «минимальным» Qst, предложенным Релетфордом (1994) [35]. Однако представление о том, что здесь принимается минимальное значение Qst, основывается на том, что h 2 всегда меньше c , что, как отмечалось ранее, является антиконсервативным. Фактически, генетическая изменчивость между популяциями, вероятно, будет меньше, чем генетическая изменчивость внутри популяций для большинства фенотипов. Наш подход аналогичен предыдущим исследованиям в том, что мы также оценили Qst, приняв c = h 2 . Однако мы использовали кривые чувствительности для оценки поведения Qst в случаях, когда c < h 2 . Кроме того, мы демонстрируем, что как внутрипопуляционные, так и межпопуляционные вариации формы носа являются наследственными.
Однако мы использовали кривые чувствительности для оценки поведения Qst в случаях, когда c < h 2 . Кроме того, мы демонстрируем, что как внутрипопуляционные, так и межпопуляционные вариации формы носа являются наследственными.
Мы провели Qst – Fst сравнения по семи признакам формы носа: ширине ноздрей, ширине крыловидного основания, высоте носа, длине носового гребня, выступу кончика носа, площади внешней поверхности и площади ноздри. Мы обнаружили, что расхождение в ширине ноздрей и крыловидного основания отклоняется от нейтральных ожиданий, и что эти результаты устойчивы к предположению c = h 2 .Это говорит о том, что ширина ноздрей и крыльев крыльев, возможно, изменилась в разных популяциях из-за дивергентного отбора.
Затем мы выдвинули гипотезу о том, что расхождение этих двух характеристик обусловлено адаптацией к климату. Чтобы исследовать это, мы проверили, коррелирует ли пространственное распределение этих признаков со средней годовой температурой и влажностью. Наши результаты показывают, что ширина ноздрей сильно коррелирует с температурой и абсолютной влажностью. Положительное направление эффектов указывает на то, что более широкие носы чаще встречаются в теплом влажном климате, а более узкие носы чаще встречаются в холодно-сухом климате.Однако ширина ноздрей не связана с относительной влажностью. Это неудивительно, поскольку абсолютная влажность, вероятно, более важна для дыхания, чем относительная влажность [43]. Ширина основания крыла только слабо, если вообще коррелирует с температурой и абсолютной влажностью, предполагая, что сигнал адаптации к климату может быть специфическим для ширины ноздрей. Исследования вычислительной гидродинамики (CFD) показывают, что геометрия носа важна для его дыхательных функций [5,44].Вдыхаемый воздух достигает 90% от требуемых уровней температуры и влажности еще до того, как достигает носоглотки, что делает полость носа, особенно носовые раковины, основным кондиционирующим аппаратом в дыхательных путях [4,5].
Наши результаты показывают, что ширина ноздрей сильно коррелирует с температурой и абсолютной влажностью. Положительное направление эффектов указывает на то, что более широкие носы чаще встречаются в теплом влажном климате, а более узкие носы чаще встречаются в холодно-сухом климате.Однако ширина ноздрей не связана с относительной влажностью. Это неудивительно, поскольку абсолютная влажность, вероятно, более важна для дыхания, чем относительная влажность [43]. Ширина основания крыла только слабо, если вообще коррелирует с температурой и абсолютной влажностью, предполагая, что сигнал адаптации к климату может быть специфическим для ширины ноздрей. Исследования вычислительной гидродинамики (CFD) показывают, что геометрия носа важна для его дыхательных функций [5,44].Вдыхаемый воздух достигает 90% от требуемых уровней температуры и влажности еще до того, как достигает носоглотки, что делает полость носа, особенно носовые раковины, основным кондиционирующим аппаратом в дыхательных путях [4,5]. Мы также знаем, что геометрия носовых дыхательных путей влияет на скорость вдыхаемого воздуха [4,7,45]. Узкие дыхательные пути в холодно-сухом климате могут способствовать лучшему кондиционированию за счет увеличения турбулентности вдыхаемого воздуха, когда он достигает носовых раковин, тем самым облегчая контакт со слизистой оболочкой носа [5].Тем не менее, мы отмечаем, что область ноздрей не показывает необычно высокой дифференциации между популяциями, что предполагает, что не размер ноздрей, а форма может иметь функциональное значение.
Мы также знаем, что геометрия носовых дыхательных путей влияет на скорость вдыхаемого воздуха [4,7,45]. Узкие дыхательные пути в холодно-сухом климате могут способствовать лучшему кондиционированию за счет увеличения турбулентности вдыхаемого воздуха, когда он достигает носовых раковин, тем самым облегчая контакт со слизистой оболочкой носа [5].Тем не менее, мы отмечаем, что область ноздрей не показывает необычно высокой дифференциации между популяциями, что предполагает, что не размер ноздрей, а форма может иметь функциональное значение.
Другая гипотеза, касающаяся функции носа, которой уделялось меньше внимания по сравнению с гипотезой кондиционирования воздуха, — это гипотеза избирательного охлаждения мозга (SBC) [46]. Известно, что некоторые крупные млекопитающие регулируют температуру мозга в гипертермических условиях, часто вызванных длительными упражнениями [47, 48].Эта способность чаще всего встречается у млекопитающих с сетчатой сонной артерией, которая представляет собой сеть капилляров в полости под мозгом, называемой кавернозным синусом. Артериальная кровь, покидающая сетку, в конечном итоге проникает в мозг через Уиллисово кольцо. Венозная кровь, идущая обратно от слизистой оболочки носа, взаимодействует с сеткой сонной артерии в кавернозном синусе и, как полагают, охлаждает кровь, поступающую в мозг у млекопитающих засушливой зоны, таких как Oryx [49], а также у акклиматизированных к зиме животных, таких как как северный олень [50].Хотя люди не обладают такой сеткой и вместо этого имеют единственную артерию, проходящую через кавернозный синус, что заставило некоторых усомниться в роли SBC у людей [51], сторонники утверждают, что сетчатка сонной артерии не является обязательным условием, поскольку SBC присутствует у некоторых животных, у которых нет сетчатки, например у лошадей [52]. Поскольку SBC у людей очень обсуждается, роль различий в форме носа в развитии различий в этом физиологическом процессе, хотя здесь стоит упомянуть, является весьма спекулятивной.
Артериальная кровь, покидающая сетку, в конечном итоге проникает в мозг через Уиллисово кольцо. Венозная кровь, идущая обратно от слизистой оболочки носа, взаимодействует с сеткой сонной артерии в кавернозном синусе и, как полагают, охлаждает кровь, поступающую в мозг у млекопитающих засушливой зоны, таких как Oryx [49], а также у акклиматизированных к зиме животных, таких как как северный олень [50].Хотя люди не обладают такой сеткой и вместо этого имеют единственную артерию, проходящую через кавернозный синус, что заставило некоторых усомниться в роли SBC у людей [51], сторонники утверждают, что сетчатка сонной артерии не является обязательным условием, поскольку SBC присутствует у некоторых животных, у которых нет сетчатки, например у лошадей [52]. Поскольку SBC у людей очень обсуждается, роль различий в форме носа в развитии различий в этом физиологическом процессе, хотя здесь стоит упомянуть, является весьма спекулятивной.
Климат, возможно, был не единственным фактором, способствовавшим различиям в форме носа у разных популяций. Фактически, мы показываем, что температура слабо коррелирует с шириной ноздрей, особенно по сравнению с корреляцией между пигментацией кожи и УФ-В. Что же тогда может быть селективным агентом, влияющим на изменение формы носа? Ранее мы упоминали, что все аспекты формы носа, изучаемые здесь, являются сексуально диморфными, что вызывает ряд вопросов. Почему существует этот половой диморфизм? Является ли это побочным продуктом циркулирующих гормонов, приводящим к различиям в росте и развитии в раннем взрослом возрасте, или он выполняет адаптивную функцию, например, сигнализирует о сексе другим мужчинам и женщинам? Половой отбор, вероятно, сыграл важную роль в эволюции человека, о чем свидетельствует наличие полового диморфизма во многих физических чертах (например,g., рост, соотношение талии и бедер, волосы на лице и грудь, и это лишь некоторые из них). Могли ли культурные различия в восприятии доминирования и привлекательности сыграть роль в расхождении формы носа [53]? Могли ли эти представления возникнуть для выбора партнеров, которые сигнализируют об адаптации к местной среде? В самом деле, экологический отбор и половой отбор могут усиливать друг друга, ускоряя фенотипическое расхождение между популяциями, несмотря на продолжающийся поток генов [54].
Фактически, мы показываем, что температура слабо коррелирует с шириной ноздрей, особенно по сравнению с корреляцией между пигментацией кожи и УФ-В. Что же тогда может быть селективным агентом, влияющим на изменение формы носа? Ранее мы упоминали, что все аспекты формы носа, изучаемые здесь, являются сексуально диморфными, что вызывает ряд вопросов. Почему существует этот половой диморфизм? Является ли это побочным продуктом циркулирующих гормонов, приводящим к различиям в росте и развитии в раннем взрослом возрасте, или он выполняет адаптивную функцию, например, сигнализирует о сексе другим мужчинам и женщинам? Половой отбор, вероятно, сыграл важную роль в эволюции человека, о чем свидетельствует наличие полового диморфизма во многих физических чертах (например,g., рост, соотношение талии и бедер, волосы на лице и грудь, и это лишь некоторые из них). Могли ли культурные различия в восприятии доминирования и привлекательности сыграть роль в расхождении формы носа [53]? Могли ли эти представления возникнуть для выбора партнеров, которые сигнализируют об адаптации к местной среде? В самом деле, экологический отбор и половой отбор могут усиливать друг друга, ускоряя фенотипическое расхождение между популяциями, несмотря на продолжающийся поток генов [54]. Это интересные направления исследований, которые необходимо рассмотреть, чтобы составить более полную картину эволюции человеческого носа.
Это интересные направления исследований, которые необходимо рассмотреть, чтобы составить более полную картину эволюции человеческого носа.
Исследование эволюции формы носа в связи с адаптацией к климату, хотя и интересно с антропологической точки зрения, также имеет медицинское значение. Поскольку люди становятся все более глобальным сообществом, изучение местной адаптации становится все более важным для понимания рисков для здоровья, связанных с жизнью в «чужом» климате. Очевидными примерами таких рисков для здоровья являются повышенный риск солнечных ожогов, рака кожи и дефицита фолиевой кислоты у людей со светлой кожей, подвергающихся высокому воздействию УФ-В, а также низкий вес при рождении и хроническая горная болезнь, связанная с гипоксией на больших высотах [20,55].Влияет ли морфология внешнего носа или внутренней носовой полости на риск респираторных заболеваний в различных климатических условиях? Пока сложно сказать. Хотя наши результаты подтверждают идею о том, что различия в аспектах формы носа могли развиваться в разных популяциях в результате давления отбора, связанного с климатом, это ранее было продемонстрировано с использованием краниометрических данных [2,12,13,41,42] Отметим, что сигнал об адаптации к климату не очень сильный, особенно по сравнению с пигментацией кожи. Это могло быть связано с более слабым давлением отбора или отбором по изменчивости насаждений, а также из-за представленной здесь разреженной выборки популяций, что является ограничением данного исследования. Эти результаты необходимо будет воспроизвести на более широком наборе групп населения. Мы ожидаем, что исследования, включающие различные группы населения, долгое время жившие в различных средах, таких как тропики, пустыни и приполярные регионы, хорошо восполнят пробелы. Особенно полезным было бы представление населения из высокогорных регионов, таких как жители Анд, тибетцы и эфиопы, которым приходится не только справляться со стрессом из-за холодного и сухого климата, но и из-за низкого уровня кислорода в атмосфере [56].Также было бы информативно изучить нечеловеческих приматов в этом контексте, которые населяют самые разные страны и демонстрируют обширные различия в морфологии носа. Наконец, будущие исследования также должны быть сосредоточены на исследованиях общегеномных ассоциаций (GWAS) для выявления вариантов, влияющих на форму носа.
Это могло быть связано с более слабым давлением отбора или отбором по изменчивости насаждений, а также из-за представленной здесь разреженной выборки популяций, что является ограничением данного исследования. Эти результаты необходимо будет воспроизвести на более широком наборе групп населения. Мы ожидаем, что исследования, включающие различные группы населения, долгое время жившие в различных средах, таких как тропики, пустыни и приполярные регионы, хорошо восполнят пробелы. Особенно полезным было бы представление населения из высокогорных регионов, таких как жители Анд, тибетцы и эфиопы, которым приходится не только справляться со стрессом из-за холодного и сухого климата, но и из-за низкого уровня кислорода в атмосфере [56].Также было бы информативно изучить нечеловеческих приматов в этом контексте, которые населяют самые разные страны и демонстрируют обширные различия в морфологии носа. Наконец, будущие исследования также должны быть сосредоточены на исследованиях общегеномных ассоциаций (GWAS) для выявления вариантов, влияющих на форму носа. С ростом интереса к идентификации локусов, связанных с формой лица, некоторые GWAS недавно идентифицировали ряд локусов формы носа [57-60]. Генетическая изменчивость в этих локусах будет информативной о природе отбора, а также для определения времени событий отбора.
С ростом интереса к идентификации локусов, связанных с формой лица, некоторые GWAS недавно идентифицировали ряд локусов формы носа [57-60]. Генетическая изменчивость в этих локусах будет информативной о природе отбора, а также для определения времени событий отбора.
Материалы и методы
Заявление об этике
Сбор данных проводился с информированного согласия участников и с одобрения институциональных наблюдательных советов (IRB) в Государственном университете Пенсильвании (IRB № 32341, IRB № 45727, IRB № 44929) и в Университете Иллинойса в Урбана-Шампейн ( IRB № 13103). Все участники предоставили письменное информированное согласие.
Набор участников и описание данных
Данные, используемые в этой статье, являются частью более крупного набора данных, который был собран в различных местах в ходе исследований, проведенных в Университете штата Пенсильвания и в Университете Иллинойса в Урбана-Шампейн.Большинство этих данных было собрано в США в Государственном университете Пенсильвании, на Всемирном фестивале науки в Нью-Йорке и в Университете Иллинойса в Урбана-Шампейн. Сбор данных в Европе происходил в Дублине (Ирландия), в Риме (Италия), в Варшаве (Польша) и в Порту (Португалия). Данные Кабо-Верде были собраны в сотрудничестве с доктором Сандрой Белеза, как описано в Beleza et al . [61]. Все данные были собраны на основе информированного согласия участников с одобрения институциональных наблюдательных советов Университета штата Пенсильвания, Университета Иллинойса в Урбана-Шампейн и Национального этического комитета медицинских исследований Кабо-Верде.
Сбор данных в Европе происходил в Дублине (Ирландия), в Риме (Италия), в Варшаве (Польша) и в Порту (Португалия). Данные Кабо-Верде были собраны в сотрудничестве с доктором Сандрой Белеза, как описано в Beleza et al . [61]. Все данные были собраны на основе информированного согласия участников с одобрения институциональных наблюдательных советов Университета штата Пенсильвания, Университета Иллинойса в Урбана-Шампейн и Национального этического комитета медицинских исследований Кабо-Верде.
Собранные фенотипические данные включают трехмерные (3D) изображения, пигментацию кожи, рост стоя и массу тела. Трехмерные изображения участников были получены с использованием системы 3dMD ( 3dMD System , Atlanta , GA ). Пигментацию кожи измеряли с помощью индекса меланина с помощью Derma Spectrometer ( Cortex Technology , Hadsund , Дания ). Индекс меланина определяется как 100 x log 10 (коэффициент отражения 1 / процент при 650 нМ) [17]. Мы сняли три показателя меланина с внутренней стороны левой и правой руки и использовали среднее значение шести измерений для каждого человека. Статистическое тестирование было выполнено с использованием инверсии индекса меланина, поскольку он имеет более нормальное распределение (S7 Рис), но использование нетрансформированного индекса меланина дает аналогичные результаты и не влияет на наши выводы. Демографическая информация была собрана с помощью анкет и включала самооценку происхождения и пола, место рождения участника, места рождения родителей, бабушек и дедушек, а также места, где участник вырос.
Мы сняли три показателя меланина с внутренней стороны левой и правой руки и использовали среднее значение шести измерений для каждого человека. Статистическое тестирование было выполнено с использованием инверсии индекса меланина, поскольку он имеет более нормальное распределение (S7 Рис), но использование нетрансформированного индекса меланина дает аналогичные результаты и не влияет на наши выводы. Демографическая информация была собрана с помощью анкет и включала самооценку происхождения и пола, место рождения участника, места рождения родителей, бабушек и дедушек, а также места, где участник вырос.
Общее количество участников — 4 257 человек. Из них мы отобрали 2637 человек на основе генетического происхождения и доступности фенотипических и ковариантных данных, как описано в следующих разделах. Сводная информация о размерах выборок, использованных в различных анализах, представлена в таблице 5. Для более подробного и точного представления обратитесь к таблице Excel в дополнительных материалах (файл S1), в которой представлены данные на индивидуальном уровне для всех образцов, используемых для создания результаты, представленные здесь.
Оценка генетического происхождения
Генотипирование и контроль качества.
Из 4 257 участников у нас были данные о генотипе 3746 человек. Поскольку данные собирались в рамках различных исследований в течение длительного периода времени, образцы были генотипированы на четырех различных наборах генотипов, а именно на чипе Illumina HumanHap300v1 BeadChip, Illumina Infinium HD Human 1M-Duo Beadarray ( Illumina , Inc ). ., San Diego , CA ) и массивы 23andMe v3 и v4 ( 23andMe Mountainview , CA) .Чтобы выбрать людей на основе генетического происхождения, нам нужно было объединить генотипы с разных платформ в единую базу данных. Мы предприняли несколько шагов, чтобы гарантировать отсутствие пакетных эффектов из-за платформы генотипирования, которые потенциально могут привести к ошибкам в оценке происхождения и вычислении Fst. Во-первых, мы удалили SNP, которые отсутствовали на всех четырех платформах, чтобы гарантировать отсутствие систематических различий из-за отсутствия. Во-вторых, мы провели контроль качества отдельно для каждой платформы, удалив людей и SNP с уровнем генотипирования менее 90%.Наконец, мы удалили палиндромные (AT / GC) SNP и перекодировали данные в формате Illumina A / B [62], чтобы гарантировать, что различия в цепях между платформами не являются проблемой. Это в достаточной мере устраняет любые потенциальные эффекты группировки из-за платформы, о чем свидетельствует наблюдение, что на графике многомерного масштабирования люди группируются по родословной, а не по платформе (S1, рис.). На этом этапе генотипы разных платформ были объединены в единый набор данных. Далее мы удалили неавтосомные SNP из объединенного набора данных и отсекли неравновесие по сцеплению (с использованием парного отсечения r 2 , равного 0.5). Это привело к 118 420 аутосомным SNP. Связанные лица были идентифицированы с использованием анализа личности по состоянию (IBS) (IBS> 0,8). Лица были удалены, чтобы минимизировать родство и максимизировать размер выборки (например, в случае квартета мать-отец-братья и сестры, братья и сестры были удалены).
Во-вторых, мы провели контроль качества отдельно для каждой платформы, удалив людей и SNP с уровнем генотипирования менее 90%.Наконец, мы удалили палиндромные (AT / GC) SNP и перекодировали данные в формате Illumina A / B [62], чтобы гарантировать, что различия в цепях между платформами не являются проблемой. Это в достаточной мере устраняет любые потенциальные эффекты группировки из-за платформы, о чем свидетельствует наблюдение, что на графике многомерного масштабирования люди группируются по родословной, а не по платформе (S1, рис.). На этом этапе генотипы разных платформ были объединены в единый набор данных. Далее мы удалили неавтосомные SNP из объединенного набора данных и отсекли неравновесие по сцеплению (с использованием парного отсечения r 2 , равного 0.5). Это привело к 118 420 аутосомным SNP. Связанные лица были идентифицированы с использованием анализа личности по состоянию (IBS) (IBS> 0,8). Лица были удалены, чтобы минимизировать родство и максимизировать размер выборки (например, в случае квартета мать-отец-братья и сестры, братья и сестры были удалены). Эти этапы контроля качества были выполнены в Plink 1.9 [63,64].
Эти этапы контроля качества были выполнены в Plink 1.9 [63,64].
Оценка геномного происхождения и анализ основных компонентов.
Мы оценили генетическое происхождение всех людей с доступными данными о генотипах (N = 3 746), используя неконтролируемый подход кластеризации в ADMIXTURE.Чтобы помочь с кластеризацией и интерпретацией полученных кластеров, мы объединили этот набор данных с генотипами из набора данных HapMap для тех же SNP (популяции включали: YRI, LWK, CEU, TSI, CHB, CHD, JPT, GIH, MEX; N = 988). Мы запустили ADMIXTURE для трех значений k (k = 5, 6, 7) и визуально определили, что оптимальная схема кластеризации будет k = 6 для наших целей. Для дальнейшей визуализации мелкомасштабной структуры мы провели неконтролируемый анализ главных компонентов (PCA) данных генотипа в Plink 1.9 [64]. Комбинация фильтров на основе результатов ADMIXTURE и оценок PCA позволила нам выбрать людей из конкретных представляющих интерес популяций. Они описаны ниже отдельно для каждого анализа.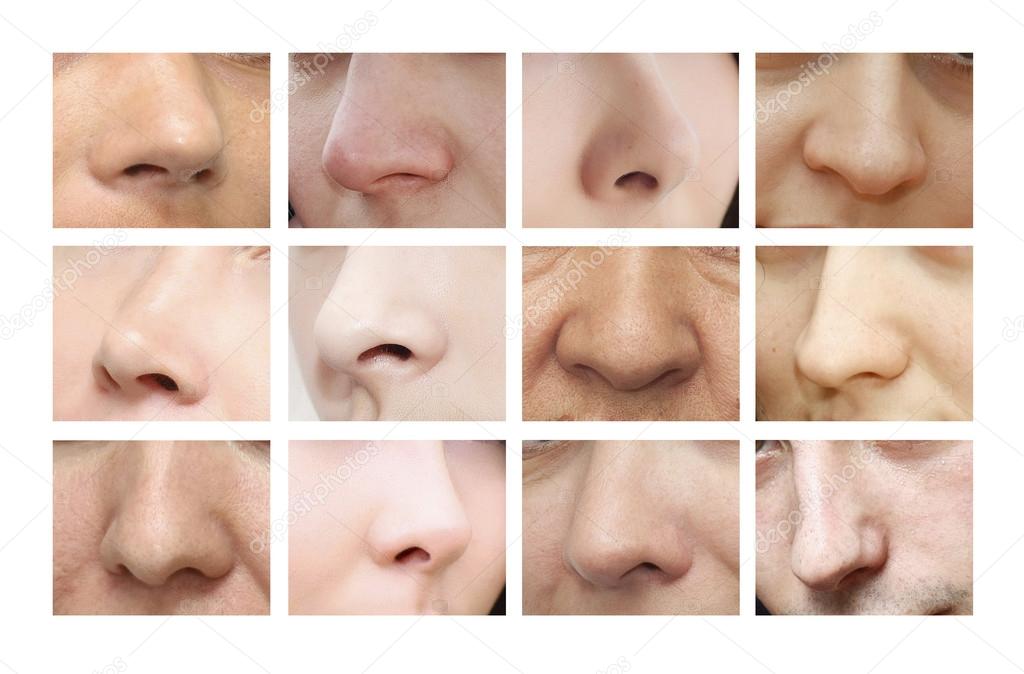
Qst – Fst анализы
Подвыборка лиц.
Для анализа Qst – Fst мы были заинтересованы в отборе людей с родословной в основном из одной из четырех популяций: i) североевропейских, ii) западноафриканских, iii) восточноазиатских и iv) южноазиатских.Во многом это было основано на генетическом происхождении, рассчитанном на основе плотных данных генотипа. Тем не менее, для некоторых людей (см. Североевропейский ниже), информация о происхождении, подтвержденная самими собой, подтвержденная через место рождения всех четырех бабушек и дедушек, использовалась там, где данные о генотипе были недоступны.
Западная Африка (N = 40) : Мы идентифицировали 40 человек, у которых было около 90% или более родословной из кластера 1. ADMIXTURE 1. Кластер 1 ADMIXTURE представляет собой предков из Западной Африки, учитывая, что люди из HapMap YRI (йоруба в Ибадане, Нигерия) происходят от большинства своих предков из кластера 1 ADMIXTURE (рис. 8).Эти 40 человек были включены в группу населения «Западная Африка».
Рис. 8. ДОБАВЛЕНИЕ Пропорции предков (k = 6) генотипированных особей, используемых в анализах Qst – Fst.
Их пропорции ADMIXTURE сравниваются с образцами из набора данных HapMap. Каждая вертикальная полоса на панелях индивидуальна, а цвета представляют долю происхождения, полученную от каждого из 6 кластеров (k = 6). На каждой панели образцы HapMap расположены слева с трехбуквенным сокращением населения, из которого они происходят (например,г. GIH относится к гуджаратским индейцам из Хьюстона, перечисленным под ними. Выборки из нашего набора данных расположены справа от выборок HapMap с обозначением их населения (например, Южная Азия) под ними.
https://doi.org/10.1371/journal.pgen.1006616.g008
Восточноазиатский (N = 127) : Сравнение с HapMap CHB (ханьский китайский в Пекине, Китай), HapMap CHD (китайский в столичном Денвере , Колорадо) и HapMap JPT (японский язык в Токио, Япония) показывает, что кластер 4 представляет генетическое происхождение из Восточной Азии (рис. 8).Мы отобрали 166 человек, у которых было 90% или более родословной из кластера 4 ADMIXTURE, для включения в группу «восточноазиатского» населения (рис. 8). Мы дополнительно изучили распределение графиков PCA и удалили шесть человек, которые сгруппировались далеко от образцов CHB, CHD и JPT. Кластеризация оставшихся 160 человек вокруг образцов CHB, CHD и JPT показана на S2 Рис. Из них у нас были высококачественные 3D-изображения и полные ковариативные данные для 127 человек, которые затем использовались в нашем анализе.
8).Мы отобрали 166 человек, у которых было 90% или более родословной из кластера 4 ADMIXTURE, для включения в группу «восточноазиатского» населения (рис. 8). Мы дополнительно изучили распределение графиков PCA и удалили шесть человек, которые сгруппировались далеко от образцов CHB, CHD и JPT. Кластеризация оставшихся 160 человек вокруг образцов CHB, CHD и JPT показана на S2 Рис. Из них у нас были высококачественные 3D-изображения и полные ковариативные данные для 127 человек, которые затем использовались в нашем анализе.
Южная Азия (N = 73) : 98 человек в нашем наборе данных имели более 60% предков из кластера 2 ADMIXTURE, который, по-видимому, представляет кластер Южной Азии (рис. 8). Эти 98 человек были добавлены к группе населения «Южная Азия». Хотя отсечение здесь кажется либеральным по сравнению с тем, что используется для других популяций, оно предназначено только в качестве приблизительного ориентира и было выбрано путем сравнения с пропорциями предков в образцах HapMap GIH (гуджаратские индейцы в Хьюстоне, Техас) (рис. 8) .Тем не менее, мы изучили распределение 98 человек в выборках GIH (гуджаратские индейцы в Хьюстоне, штат Техас), используя графики PCA (S2 рис.), И заметили, что шесть человек, по-видимому, упали далеко от основного кластера, которые впоследствии были удалены. Из оставшихся у нас были высококачественные 3D-изображения и полные ковариативные данные для 73 человек, которые использовались для дальнейшего анализа.
8) .Тем не менее, мы изучили распределение 98 человек в выборках GIH (гуджаратские индейцы в Хьюстоне, штат Техас), используя графики PCA (S2 рис.), И заметили, что шесть человек, по-видимому, упали далеко от основного кластера, которые впоследствии были удалены. Из оставшихся у нас были высококачественные 3D-изображения и полные ковариативные данные для 73 человек, которые использовались для дальнейшего анализа.
Северная Европа (N = 236) : В нашем полном наборе данных было большое количество людей европейского происхождения из разных регионов Европы.Чтобы минимизировать структуру в Европе, мы оставили только лиц североевропейского происхождения. Таким образом, мы ограничили анализ европейцами, собранными в Ирландии (N = 151) и Польше (N = 85), которые сообщили, что предки всех четверых их бабушек и дедушек были из региона отбора. Более подробная информация об этих образцах описана в Candille et al . [65]. Из этого набора у нас были генотипы 73 женщин (ирландцы = 37, поляки = 36), но ни одного для мужчин.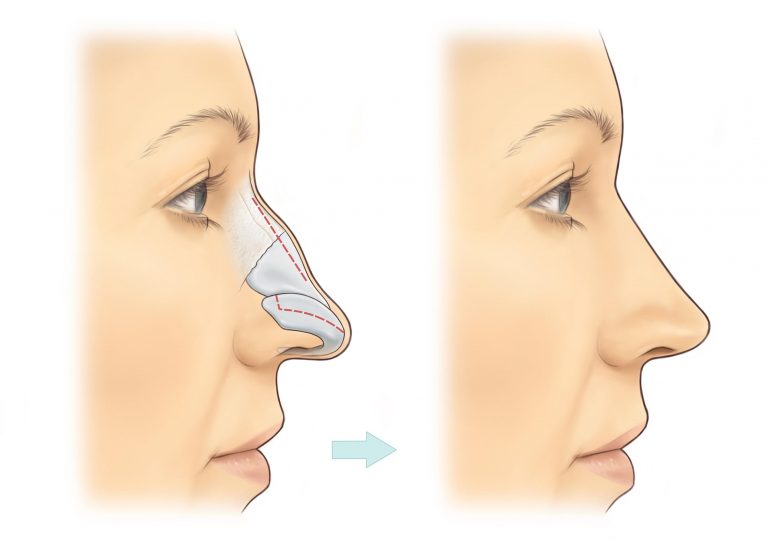 Это причина того, что только самки были включены в фенотип-климатический анализ, где генотипы использовались для построения матрицы генетических отношений.Результаты ADMIXTURE для этих женщин показаны на рис. 8 вместе с результатами выборок CEU (жители Юты северного и западноевропейского происхождения из коллекции CEPH) и TSI (тосканцы из Италии).
Это причина того, что только самки были включены в фенотип-климатический анализ, где генотипы использовались для построения матрицы генетических отношений.Результаты ADMIXTURE для этих женщин показаны на рис. 8 вместе с результатами выборок CEU (жители Юты северного и западноевропейского происхождения из коллекции CEPH) и TSI (тосканцы из Италии).
Расчет Qst.
Оценка компонентов дисперсии, и, была проведена с использованием линейной модели, в которой группа населения рассматривалась как фиксированный эффект с полом, возрастом и ИМТ в качестве ковариант. Если MSB — средний квадрат среди популяций, MSW — средний квадрат внутри популяций, n i — это количество особей в -й популяциях, а a — это количество популяций, и их можно оценить следующим образом [10,66,67]: (3) (4) (5)
Fst расчет.
Fst Райта измеряет генетическое расхождение между двумя или более популяциями. Мы оценили Fst в четырех популяциях для LD-сокращенного набора из 118 420 аутосомных SNP, описанного выше, с использованием шкалы Вейра и Кокерхема θ [68]. Распределение геномного Fst показано на рис. S3
Распределение геномного Fst показано на рис. S3
Qst – Fst сравнение.
Мы использовали непараметрический подход начальной загрузки, чтобы проверить, действительно ли Qst фенотипа значительно больше, чем Fst. Это было сделано путем вычисления Qst из подвыборок, сгенерированных случайным выбором лиц с замещением, в пределах популяции и пола так, чтобы количество мужчин и женщин в каждой популяции оставалось неизменным.Значения Fst моделировались путем случайной выборки из Fst 118 420 SNP (S3, рис.). Мы сгенерировали 10 000 таких псевдовыборок и для каждого смоделированного значения Qst и Fst вычислили разность Qst – Fst. Высокие значения Qst – Fst указывают на дивергентный выбор, и эмпирическое значение p для этого теста было определено путем расчета доли значений Qst – Fst, которые меньше нуля.
Оценка и
cОценка.
Для расчета мы определили 1825 неродственных особей преимущественно европейского происхождения, у которых более 80% своей комбинированной генетической родословной произошло из кластера 3 и кластера 6 на основе результатов ADMIXTURE (рис. 9).Это пороговое значение было выбрано на основе сравнения с генетическими предками людей CEU (жители штата Юта с северным и западноевропейским происхождением из коллекции CEPH) и TSI (тосканы в Италии) из набора данных HapMap (рис. 9). Мы использовали GCTA для расчета полногеномной матрицы родства для 1825 человек из 118 420 аутосомных SNP, которые перекрывались на всех четырех платформах. Из 1825 человек европейского происхождения мы измерили рост 1825 человек, измерили индекс меланина 1231 и сделали 3D-фотографии 1718 человек.На основе этих данных мы использовали GCTA для оценки доли фенотипической дисперсии, объясняемой генотипированными SNP ( h g 2 ) для аспектов роста, пигментации кожи и формы носа. Четыре основных генетических ПК были включены в модель как ковариаты для корректировки структуры популяции, которая может привести к завышению оценок наследуемости [18,69,70]. Также в модель были включены пол, возраст и ИМТ.
9).Это пороговое значение было выбрано на основе сравнения с генетическими предками людей CEU (жители штата Юта с северным и западноевропейским происхождением из коллекции CEPH) и TSI (тосканы в Италии) из набора данных HapMap (рис. 9). Мы использовали GCTA для расчета полногеномной матрицы родства для 1825 человек из 118 420 аутосомных SNP, которые перекрывались на всех четырех платформах. Из 1825 человек европейского происхождения мы измерили рост 1825 человек, измерили индекс меланина 1231 и сделали 3D-фотографии 1718 человек.На основе этих данных мы использовали GCTA для оценки доли фенотипической дисперсии, объясняемой генотипированными SNP ( h g 2 ) для аспектов роста, пигментации кожи и формы носа. Четыре основных генетических ПК были включены в модель как ковариаты для корректировки структуры популяции, которая может привести к завышению оценок наследуемости [18,69,70]. Также в модель были включены пол, возраст и ИМТ.
Рис. 9. ПРИМЕСЬ пропорции предков (k = 6) 1825 человек преимущественно европейского происхождения, которые были использованы для оценки h g 2 .
9. ПРИМЕСЬ пропорции предков (k = 6) 1825 человек преимущественно европейского происхождения, которые были использованы для оценки h g 2 .
Каждая полоса представляет человека, а разные цвета представляют пропорции ПРИМЕСИ из шести кластеров (k = 6). Пропорции ADMIXTURE для европейских популяций HapMap (CEU и TSI) показаны слева для сравнения.
https://doi.org/10.1371/journal.pgen.1006616.g009
Оценка c.
Мы оценили долю фенотипической дисперсии между популяциями из-за аддитивных генетических эффектов ( c ) в смешанной популяции из Кабо-Верде, которые произошли в основном от W.Африканцы и европейцы (рис. 10), используя подход, описанный в Zaitlen et al . [40]. 697 жителей Кабо-Верде были генотипированы на Illumina Infinium HD Human 1M-Duo Beadarray ( Illumina , Inc ., San Diego , CA ) в общей сложности 1 016 423 SNP. Это было сокращено до 623 625 аутосомных SNP после удаления палиндромных SNP, SNP, которые не перекрывались с набором данных HapMap, и других SNP на основе фильтров, описанных в Beleza et al . [17]. У нас были высококачественные 3D-изображения и полные ковариативные данные для 409 из 697 жителей Кабо-Верде, которые были генотипированы. Для этих людей мы рассчитали местное происхождение по 623 625 аутосомным SNP с LAMP-LD [71], используя поэтапные генотипы из образцов HapMap CEU и YRI в качестве эталона. Усреднение местного происхождения по всем SNP дает оценку глобального происхождения, которая сильно коррелирует с оценками, полученными из ADMIXTURE с использованием k = 2 (r 2 ∼ 1) (S4 Рис.). Мы использовали GCTA для расчета матрицы родства на основе местного происхождения и для оценки доли фенотипической дисперсии, объясняемой местным происхождением ( h y 2 ).Глобальное происхождение, рассчитанное на основе среднего местного происхождения по всем SNP, было включено в качестве ковариаты для корректировки стратификации предков и любых переменных окружающей среды, которые могут совпадать с родословной [16,72].
[17]. У нас были высококачественные 3D-изображения и полные ковариативные данные для 409 из 697 жителей Кабо-Верде, которые были генотипированы. Для этих людей мы рассчитали местное происхождение по 623 625 аутосомным SNP с LAMP-LD [71], используя поэтапные генотипы из образцов HapMap CEU и YRI в качестве эталона. Усреднение местного происхождения по всем SNP дает оценку глобального происхождения, которая сильно коррелирует с оценками, полученными из ADMIXTURE с использованием k = 2 (r 2 ∼ 1) (S4 Рис.). Мы использовали GCTA для расчета матрицы родства на основе местного происхождения и для оценки доли фенотипической дисперсии, объясняемой местным происхождением ( h y 2 ).Глобальное происхождение, рассчитанное на основе среднего местного происхождения по всем SNP, было включено в качестве ковариаты для корректировки стратификации предков и любых переменных окружающей среды, которые могут совпадать с родословной [16,72]. Пол, возраст и ИМТ также были включены в качестве ковариант.
Пол, возраст и ИМТ также были включены в качестве ковариант.
Рис. 10. ПРИМЕСЬ пропорции предков (k = 6) 409 жителей Кабо-Верде, которые были использованы для оценки c .
Каждая полоса представляет человека, а разные цвета представляют пропорции ПРИМЕСИ из шести кластеров (k = 6).
https://doi.org/10.1371/journal.pgen.1006616.g010
Фенотипо-климатические ассоциации
Тестирование на локальную адаптацию.
Основное внимание здесь было уделено ответу на вопрос: показывает ли изменение формы носа между географически удаленными популяциями сигналы местной адаптации? Чтобы ответить на этот вопрос, нам нужно было приписать каждому человеку климатическую ценность, которая может служить представителем среды их предков. По этой причине мы выбрали людей, использованных в анализе Qst – Fst, которые сообщили по крайней мере о месте рождения одного из родителей.Они были дополнительно отфильтрованы для удержания лиц, чьи места рождения родителей совпадали с их континентальным происхождением, например, лица из Восточной Азии, чьи родители родились в Соединенных Штатах, были удалены.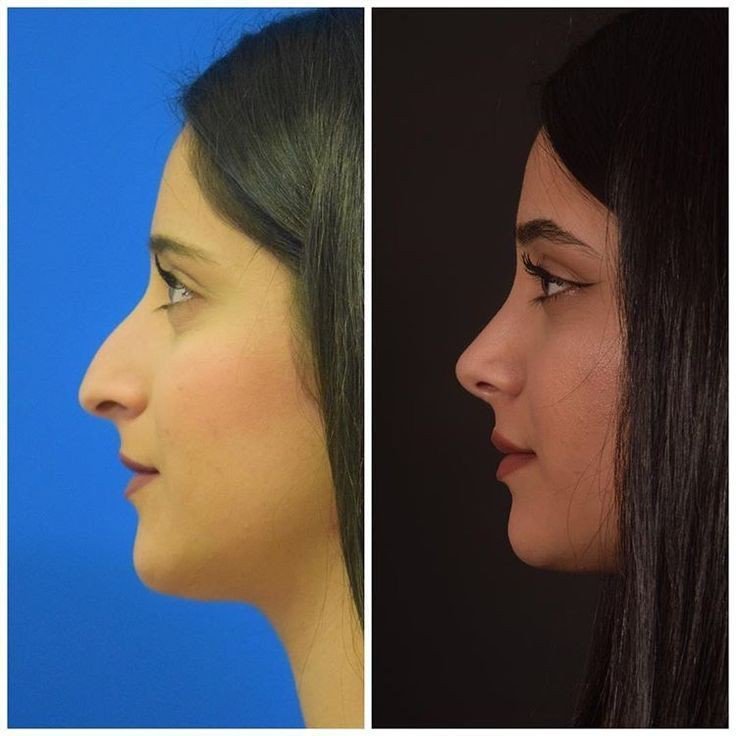 Наконец, мы ограничили анализ женщинами, поскольку генетические данные не были доступны для мужчин, отобранных в Ирландии и Польше. В результате получено 140 женщин с генетическими, фенотипическими и географическими данными. Мы изобразили географическое распределение родителей этих людей на рис. 6 линией, соединяющей двух родителей каждого человека.Одиночная точка означает, что родители родились в одном месте. Эти местоположения были геокодированы для получения широты и долготы в десятичных градусах с использованием пакета ggmap в R [73,74].
Наконец, мы ограничили анализ женщинами, поскольку генетические данные не были доступны для мужчин, отобранных в Ирландии и Польше. В результате получено 140 женщин с генетическими, фенотипическими и географическими данными. Мы изобразили географическое распределение родителей этих людей на рис. 6 линией, соединяющей двух родителей каждого человека.Одиночная точка означает, что родители родились в одном месте. Эти местоположения были геокодированы для получения широты и долготы в десятичных градусах с использованием пакета ggmap в R [73,74].
Климатические данные с координатной привязкой были получены в системе координат широта / долгота. Климатические сетки со среднемесячными значениями температуры (в градусах Цельсия) и давления пара (в гектопаскалях или гПа) с разрешением 0,5 ° за период с 1901 по 2015 год были получены из The Climate Research Unit TS v3.24 Dataset (https://crudata.uea.ac.uk/cru/data/hrg/cru_ts_3.24/) [75]. Данные приземного ультрафиолетового излучения B (UVB) (выраженные в Дж / м 2 ) за период между 1996 и 2005 годами, полученные командой Earth Probe Total Ozone Mapping Spectrometer (EP / TOMS) с разрешением 0,5 °, были загружены с веб-сайта Центра данных и информационных услуг Годдарда (GES DISC) (https://disc. gsfc.nasa.gov/) [76]. Сетки относительной и абсолютной влажности были получены на основе температуры и давления пара, как описано в Maddux et al .(2016) [43]. Для каждой из четырех климатических переменных (UVB, температура, абсолютная влажность и относительная влажность) была построена единая сетка путем усреднения по всем месяцам в пределах периода времени, чтобы представить долгосрочный климат. Пакеты R RnetCDF [77] и Raster [78] использовались для чтения данных с координатной привязкой и для извлечения значений климата в каждом месте. Мы использовали значения климата в месте рождения родителей, чтобы представить климат предков для каждого человека. Если было известно место рождения обоих родителей, использовались средние значения климата, полученные в двух местах.Оба родителя обычно происходят из близлежащих регионов, по крайней мере, в континентальном масштабе (рис. 6), и значения климата для родителей очень похожи (S5 рис.).
gsfc.nasa.gov/) [76]. Сетки относительной и абсолютной влажности были получены на основе температуры и давления пара, как описано в Maddux et al .(2016) [43]. Для каждой из четырех климатических переменных (UVB, температура, абсолютная влажность и относительная влажность) была построена единая сетка путем усреднения по всем месяцам в пределах периода времени, чтобы представить долгосрочный климат. Пакеты R RnetCDF [77] и Raster [78] использовались для чтения данных с координатной привязкой и для извлечения значений климата в каждом месте. Мы использовали значения климата в месте рождения родителей, чтобы представить климат предков для каждого человека. Если было известно место рождения обоих родителей, использовались средние значения климата, полученные в двух местах.Оба родителя обычно происходят из близлежащих регионов, по крайней мере, в континентальном масштабе (рис. 6), и значения климата для родителей очень похожи (S5 рис.).
Мы использовали линейную смешанную модель для проверки взаимосвязи между фенотипом и климатическими переменными с поправкой на возраст и ИМТ. Мы приняли во внимание отсутствие независимости между наблюдениями из-за генетического сходства, позволив коррелировать наблюдения в соответствии с основанной на SNP полногеномной матрице родства. В матричной записи линейная модель принимает следующий вид:
(6)
Где β — фиксированные эффекты, а ε — член случайной ошибки.Таким образом, ε ∼ N (0, Gσ e ), где G — полногеномная матрица родства, которая была построена из генотипов по 118 420 SNP в GCTA [37]. Наклоны оценивались с использованием метода максимального правдоподобия, реализованного в пакете spaMM в R [74,79]. P-значения для каждого наклона были получены из отношения правдоподобия между полной и сокращенной моделями (подгонка путем удаления каждого предиктора климата).
Мы приняли во внимание отсутствие независимости между наблюдениями из-за генетического сходства, позволив коррелировать наблюдения в соответствии с основанной на SNP полногеномной матрице родства. В матричной записи линейная модель принимает следующий вид:
(6)
Где β — фиксированные эффекты, а ε — член случайной ошибки.Таким образом, ε ∼ N (0, Gσ e ), где G — полногеномная матрица родства, которая была построена из генотипов по 118 420 SNP в GCTA [37]. Наклоны оценивались с использованием метода максимального правдоподобия, реализованного в пакете spaMM в R [74,79]. P-значения для каждого наклона были получены из отношения правдоподобия между полной и сокращенной моделями (подгонка путем удаления каждого предиктора климата).
Морфометрические измерения
Обработка изображений.
Мы сделали трехмерные изображения лиц участников с высоким разрешением с помощью системы 3dMD Face ( 3dMD Atlanta , GA ). Пять ориентиров для позиционирования (два на внутреннем углу глаз, два на внешних углах рта и один на кончике носа) были нанесены на каждое лицо, чтобы установить ориентацию лица. Пространственно плотная сетка из 7 150 квази-ориентиров (QL) была нанесена на каждое изображение и его отражение. В результате этого отображения вершины и треугольники гомологичны для всех людей [80].Обобщенное наложение Прокруста [24] было выполнено на обоих наборах изображений (исходном и отраженном) для устранения различий в положении и ориентации. Координаты Прокруста исходного и отраженного изображений для каждого участника затем были усреднены, чтобы устранить эффекты двусторонней асимметрии. Мы выбрали область носа (см. Схему на рис. 1), которая состоит из 709 из 7 150 QL, составляющих все лицо, для дальнейшего анализа.
Пять ориентиров для позиционирования (два на внутреннем углу глаз, два на внешних углах рта и один на кончике носа) были нанесены на каждое лицо, чтобы установить ориентацию лица. Пространственно плотная сетка из 7 150 квази-ориентиров (QL) была нанесена на каждое изображение и его отражение. В результате этого отображения вершины и треугольники гомологичны для всех людей [80].Обобщенное наложение Прокруста [24] было выполнено на обоих наборах изображений (исходном и отраженном) для устранения различий в положении и ориентации. Координаты Прокруста исходного и отраженного изображений для каждого участника затем были усреднены, чтобы устранить эффекты двусторонней асимметрии. Мы выбрали область носа (см. Схему на рис. 1), которая состоит из 709 из 7 150 QL, составляющих все лицо, для дальнейшего анализа.
Линейные расстояния.
Линейные расстояния были рассчитаны с использованием трех ориентиров средней линии; я.назион (п), II. пронасале (прн), iii. subnasale (sn) и два набора парных ориентиров; iv. искривление левой и правой крыльев (al l и al r ) и v. левое и правое основание крыла (ac l и ac r ). Эти ориентиры были определены ранее [1,26], и их приблизительное положение показано на рис. 1. Два разных наблюдателя поместили каждую из этих семи ориентиров три раза в разное время на пространственно-плотной сетке шаблона QL, нанесенной на все грани.Поскольку QL гомологичны для всех людей, мы можем автоматически выбрать соответствующие ориентиры для всех людей, указав их в шаблоне QL. Это возможно, потому что любая точка на поверхности шаблона может быть выражена как функция QL с использованием барицентрических координат на треугольниках. Используя этот подход, мы получили координаты x, y и z вышеупомянутых ориентиров для каждого человека. Затем евклидовы расстояния (в мм) были рассчитаны отдельно для каждого из шести повторных размещений, которые были усреднены по повторениям, а затем усреднены между двумя наблюдателями, чтобы получить средние расстояния для каждого человека.
искривление левой и правой крыльев (al l и al r ) и v. левое и правое основание крыла (ac l и ac r ). Эти ориентиры были определены ранее [1,26], и их приблизительное положение показано на рис. 1. Два разных наблюдателя поместили каждую из этих семи ориентиров три раза в разное время на пространственно-плотной сетке шаблона QL, нанесенной на все грани.Поскольку QL гомологичны для всех людей, мы можем автоматически выбрать соответствующие ориентиры для всех людей, указав их в шаблоне QL. Это возможно, потому что любая точка на поверхности шаблона может быть выражена как функция QL с использованием барицентрических координат на треугольниках. Используя этот подход, мы получили координаты x, y и z вышеупомянутых ориентиров для каждого человека. Затем евклидовы расстояния (в мм) были рассчитаны отдельно для каждого из шести повторных размещений, которые были усреднены по повторениям, а затем усреднены между двумя наблюдателями, чтобы получить средние расстояния для каждого человека.
Область ноздри.
Чтобы измерить площадь ноздрей, наблюдатели трижды определили области вокруг ноздрей (примерное местоположение показано на рис. 1). Площадь региона рассчитывалась как общая площадь многоугольников внутри региона. Используя этот подход, мы рассчитали среднюю площадь правой и левой ноздрей (в мм 2 ) для каждой повторности. Затем это было усреднено по повторам для одного и того же наблюдателя, а затем по всем наблюдателям, чтобы получить среднюю площадь ноздрей для каждого человека.
Наружная поверхность.
Площадь внешней поверхности носа (также в мм 2 ) была рассчитана путем суммирования площадей многоугольников, ограниченных 709 QL, определяющих всю область носа (показано на рис. 1).
Ошибка наблюдателя и надежность наблюдателя.
Мы рассчитали стандартное отклонение при измерении линейных расстояний (в мм) и площади ноздрей (в мм 2 ), сделанных двумя наблюдателями для оценки степени ошибки внутри наблюдателя. Квантили для этих стандартных отклонений показаны в таблице 6. Измерения Observer2 немного более внутренне согласованы, чем Observer1. Для Observer1 выступ кончика носа был наименее точным измерением расстояния с 95% стандартных отклонений в пределах от 0,791 мм до 1,165 мм, с ошибкой для остальных расстояний значительно меньше 1 мм. Распределение ошибок для области ноздрей было одинаковым для Observer1 (95% CI: 0,914–1,930) и Observer2 (95% CI: 0,888–2,243).
Квантили для этих стандартных отклонений показаны в таблице 6. Измерения Observer2 немного более внутренне согласованы, чем Observer1. Для Observer1 выступ кончика носа был наименее точным измерением расстояния с 95% стандартных отклонений в пределах от 0,791 мм до 1,165 мм, с ошибкой для остальных расстояний значительно меньше 1 мм. Распределение ошибок для области ноздрей было одинаковым для Observer1 (95% CI: 0,914–1,930) и Observer2 (95% CI: 0,888–2,243).
Надежность между наблюдателями оценивалась с использованием коэффициента внутриклассовой корреляции (ICC).ICC для каждой меры рассчитывалась с использованием линейных смешанных моделей, где наблюдатель рассматривался как фиксированный эффект, а субъект рассматривался как случайный эффект. Это было выполнено с использованием пакета nlme в R [74,81]. За исключением выступа кончика носа (ICC = 0,819), ICC для всех других показателей были довольно высокими (ICC> 0,95). Это соответствие очевидно при сравнении средних измерений между Observer1 и Observer2 (S6 Fig)
Вспомогательная информация
S1 Рис.

Графики MDS (k = 2), показывающие кластеризацию генотипированных индивидуумов по A) платформе генотипирования и B) по родословной. В B) генотипированные индивидуумы из исследуемой группы показаны черным, в то время как образцы HapMap показаны цветом для справки. График показывает, что люди группируются по родословной, а не по платформе генотипирования. Таким образом, пакетные эффекты из-за платформы генотипирования вряд ли приведут к артефактам при выводе родословной.
https://doi.org/10.1371/journal.pgen.1006616.s001
(TIF)
S2 Рис.
Графики PCA, показывающие кластеризацию генотипированных индивидуумов по A) южноазиатскому и B) восточноазиатскому происхождению вокруг эталонных образцов HapMap. Цветные точки — эталонные образцы из набора данных HapMap, а серые точки — отдельные лица из нашего набора данных. A) Люди из Южной Азии показаны с образцами HapMap GIH, а B) Люди из Восточной Азии показаны с объединенной выборкой из Восточной Азии (CHB + JPT + CHD) из набора данных HapMap. Красные линии указывают на порог удаления лиц, которые, кажется, сгруппированы далеко от основного кластера. Лица, которые были удалены на основе кластеризации, показаны треугольниками, а все остальные показаны кружками.
Красные линии указывают на порог удаления лиц, которые, кажется, сгруппированы далеко от основного кластера. Лица, которые были удалены на основе кластеризации, показаны треугольниками, а все остальные показаны кружками.
https://doi.org/10.1371/journal.pgen.1006616.s002
(TIF)
S4 Рис. Глобальное происхождение, рассчитанное с помощью LAMP-LD, сильно коррелирует с глобальным происхождением, рассчитанным с помощью ADMIXTURE.
Оценки предков на основе ADMIXTURE были рассчитаны с использованием допущения k = 2. При оценке происхождения на основе LAMP-LD вычислялось среднее местное происхождение по 623 625 аутосомным SNP.
https://doi.org/10.1371/journal.pgen.1006616.s004
(TIF)
S7 Рис.
QQplots, показывающие, что остатки для A) обратного индекса меланина распределены более нормально, чем B) нетрансформированного индекса меланина. Точки представляют собой остатки от линейных моделей, в которых в качестве предикторов использовались популяция, пол, возраст и ИМТ.
https://doi.org/10.1371/journal.pgen.1006616.s007
(TIF)
Благодарности
Авторы прежде всего хотели бы поблагодарить участников исследования, без которых все это было бы невозможно.Мы благодарны команде Shriver Lab, Рипану Малхи, и членам его лаборатории за их усилия по сбору и организации данных. Мы также благодарны Сандре Белезе и Изабель Инес Араужу за их усилия по сбору и распространению данных по Кабо-Верде. Мы благодарим Ине Сай из группы Питера Класа за помощь с морфометрическими анализами. Мы также благодарим Нину Яблонски, Джорджа Чаплина, Джорджа Перри, Лорел Пирсон, Джули Уайт и Мэтью Реймхерра за полезные обсуждения аналитических аспектов статьи.Наконец, мы хотели бы поблагодарить двух анонимных рецензентов за полезную критику и комментарии к первому представлению этой рукописи.
Вклад авторов
- Концептуализация: AAZ MDS.
- Обработка данных: AAZ BCM MDS BM.
- Формальный анализ: AAZ.
- Финансирование: MDS PC CH.
- Исследование: AAZ BCM CH MDS.
- Методология: ААЗ МДС.
- Администрация проекта: МДС СН БЦМ БМ.
- Ресурсы: MDS CH PC BM.
- Программное обеспечение: PC AAZ.
- Кураторство: МДС.
- Визуализация: AAZ PC.
- Написание — черновик: AAZ.
- Написание — просмотр и редактирование: AAZ MDS PC CH BCM BM.

Ссылки
- 1. Фаркас Л.Г., Колар Дж.С., Манро И.Р.География носа: морфометрическое исследование. Эстетическая Пласт Сург. 1986; 10: 191–223. pmid: 3812136
- 2. Францискус Р.Г., Лонг Дж.С. Различия в высоте и ширине носа у человека. Am J Phys Anthropol. 1991; 85: 419–427. pmid: 1928315
- 3.
Negus VE. Введение в сравнительную анатомию носа и придаточных пазух носа. Ann R Coll Surg Engl. 1954; 15: 141–171. pmid: 13198060
- 4. Ханна Л.М., Шерер П.В. Теоретическая модель локального переноса тепла и водяного пара в дыхательных путях человека.J Biomech Eng. Американское общество инженеров-механиков; 1986; 108: 19. Pmid: 3959548
- 5. Нафтали С., Розенфельд М., Вольф М., Элад Д. Способность человеческого носа к кондиционированию воздуха. Ann Biomed Eng. 2005. 33: 545–553. pmid: 15 0
- 6. Рэнделл Ш., Бушер Р. Эффективное очищение от слизи необходимо для здоровья дыхательных путей. Am J Respir Cell Mol Biol. 2006; 35: 20–8. pmid: 16528010
- 7. Черчилль С.Е., Шакелфорд Л.Л., Георгий Дж. Н., Блэк МТ.Морфологические изменения и динамика воздушного потока в носу человека. Am J Hum Biol. 2004. 16: 625–638. pmid: 15495233
- 8.
Weiner JS. Форма носа и климат. Am J Phys Anthropol. 1954; 12: 615–618. pmid: 14350081
- 9. Роземан С.С., Уивер Т.Д. Многомерное распределение глобального краниометрического разнообразия человека. Am J Phys Anthropol. 2004. 125: 257–263. pmid: 15386236
- 10. Спитце К. Популяционная структура Daphnia obtusa: количественная генетическая и аллозимическая изменчивость.Генетика. 1993. 135: 367–374. Доступно: http://www.genetics.org/content/135/2/367 pmid: 8244001
- 11. Whitlock MC, Guillaume F. Тестирование пространственно расходящегося отбора: сравнение QST с FST. Генетика. 2009; 183: 1055–63. pmid: 19687138
- 12. Roseman CC. Выявление межрегионального разнообразия естественного отбора на современной черепной форме человека с использованием сопоставленных молекулярных и морфометрических данных. Proc Natl Acad Sci U S. A. 2004; 101: 12824–9. pmid: 15326305
- 13.Хуббе М., Ханихара Т., Харвати К. Климатические сигнатуры в морфологической дифференциации современных человеческих популяций во всем мире. Анат Рек. 2009; 292: 1720–1733.
- 14. Го Дж, Тан Дж, Ян Й, Чжоу Х, Ху С, Хашан А и др. Вариации и подписи выбора на человеческом лице. J Hum Evol. Elsevier Ltd; 2014; 75: 143–152. pmid: 25186351
- 15. Шрайвер, доктор медицины, Парра Э., Дж. Диос, С. Бонилла, Нортон Н., Джовел С. и др. Пигментация кожи, биогеографическое происхождение и составление карт примесей.Hum Genet. 2003; 112: 387–99. pmid: 12579416
- 16. Парра Э.Дж., Киттлс Ра, Шрайвер Мэриленд. Значение корреляции между цветом кожи и генетическим происхождением для биомедицинских исследований. Нат Жене. 2004; 36: S54 – S60. pmid: 15508005
- 17. Белеза С., Джонсон Н.А., Кандилль С.И., Абшер Д.М., Корам М.А., Лопес Дж. И др. Генетическая архитектура кожи и цвета глаз в смешанном афро-европейском населении. Сприц Р.А., редактор. PLoS Genet. Публичная научная библиотека; 2013; 9: e1003372.pmid: 23555287
- 18.
Ян Дж., Беньямин Б. , Макэвой Б. П., Гордон С., Хендерс А. К., Найхольт Д. Р. и др. Распространенные SNP объясняют большую часть наследуемости по росту человека. Нат Жене. Nature Publishing Group, подразделение Macmillan Publishers Limited. Все права защищены.; 2010; 42: 565–9. pmid: 20562875
- 19. Turchin MC, Chiang CWK, Palmer CD, Sankararaman S, Reich D, Hirschhorn JN. Свидетельства широко распространенного отбора вариаций стояния в Европе по SNP, связанным с ростом.Нат Жене. Nature Publishing Group, подразделение Macmillan Publishers Limited. Все права защищены.; 2012; 44: 1015–9. pmid: 227
- 20. Яблонски Н.Г. Эволюция кожи человека и цвета кожи. Анну Преподобный Антрополь. 2004. 33: 585–623.
- 21. Ламасон Р.Л., Мохидин М-АПК, Мест-Дж. Р., Вонг А.С., Нортон Х.Л., Арос М.С. и др. Предполагаемый катионообменник SLC24A5 влияет на пигментацию у рыбок данио и людей. Наука. 2005; 310: 1782–6. pmid: 16357253
- 22.Нортон Х.Л., Киттлс Р. А., Парра Э., МакКейг П., Мао Х, Ченг К. и др. Генетические доказательства конвергентной эволюции светлой кожи у европейцев и выходцев из Восточной Азии. Mol Biol Evol. 2007; 24: 710–22. pmid: 17182896
- 23. Яблонский Н.Г., Чаплин Г. Эволюция окраски кожи человека. J Hum Evol. 2000. 39: 57–106. pmid: 10896812
- 24. Gower JC. Обобщенный анализ прокрастов. Психометрика. 1975; 40: 33–51.
- 25. Клас П., Либертон Д.К., Дэниэлс К., Розана К.М., Квиллен Е.Е., Пирсон Л.Н. и др.Моделирование трехмерной формы лица по ДНК. Лукетти Д., редактор. PLoS Genet. Публичная научная библиотека; 2014; 10: e1004224. pmid: 24651127
- 26. Феррарио В.Ф., Миан Ф., Перетта Р., Розати Р., Сфорца С. Трехмерная компьютеризированная антропометрия носа: представление ориентира по сравнению с анализом поверхности. Расщелина неба и черепно-лицевой области J. 2007; 44: 278–285.
- 27.
Nucara A, Pietrafesa M, Rizzo G, Scaccianoce G. Справочник по антропометрии. Handb Anthr. 2012; 91–114.
- 28. Leinonen T, McCairns RJS, O’Hara RB, Merilä J. Сравнение Q (ST) -F (ST): эволюционное и экологическое понимание геномной гетерогенности. Nat Rev Genet. Nature Publishing Group, подразделение Macmillan Publishers Limited. Все права защищены.; 2013; 14: 179–90. pmid: 23381120
- 29. Райт С. Генетическая структура популяций. Энн Ойген. 1949; 15: 323–354.
- 30. Ланде Р. Нейтральная теория количественной генетической изменчивости в модели острова с локальным исчезновением и колонизацией.Эволюция (N Y). 1992; 46: 381–389.
- 31. Whitlock MC. Эволюционный вывод из QST. Молекулярная экология. 2008. С. 1885–1896. pmid: 18363667
- 32. Brommer JE. Куда Pst? Приближение Qst к Pst в эволюционной биологии и биологии сохранения. J Evol Biol. 2011; 24: 1160–8. pmid: 21457173
- 33.
Лейнонен Т., Кано Дж. М., Мякинен Х., Мерила Дж. Контрастные модели формы тела и нейтральное генетическое расхождение в морских и озерных популяциях трехиглой колюшки.J Evol Biol. 2006; 19: 1803–12. pmid: 17040377
- 34. Perry GH, Foll M, Grenier JC, Patin E, Nédélec Y, Pacis A и др. Адаптивное, конвергентное происхождение фенотипа пигмеев у охотников-собирателей тропических лесов Африки. Proc Natl Acad Sci U S. A. 2014; 111: E3596–603. pmid: 25136101
- 35. Релетфорд Дж. Х. Краниометрические вариации среди современных человеческих популяций. Am J Phys Anthropol. 1994; 95: 53–62. pmid: 7527996
- 36. Линч М., Уолш Б. Генетика и анализ количественных признаков.Сандерленд, Массачусетс: Sinauer Associates, Inc; 1998.
- 37. Ян Дж., Ли Ш., Годдард М.Э., Вишер ПМ. GCTA: инструмент для анализа сложных признаков в масштабе всего генома. Am J Hum Genet. 2011; 88: 76–82. pmid: 21167468
- 38.
Sabeti PC, Varilly P, Fry B, Lohmueller J, Hostetter E, Cotsapas C и др. Полногеномное обнаружение и характеристика положительного отбора в человеческих популяциях. Природа. 2007; 449: 913–918. pmid: 17943131
- 39. Дарваси А., Шифман С.Красота смеси. Нат Жене. 2005. 37: 118–119. pmid: 15678141
- 40. Зайтлен Н., Пасанюк Б., Санкарараман С., Бхатиа Дж., Чжан Дж., Гусев А. и др. Использование примеси популяции для характеристики наследуемости сложных признаков. Нат Жене. Nature Publishing Group, подразделение Macmillan Publishers Limited. Все права защищены.; 2014; 46: 1356–1362. pmid: 25383972
- 41. Yokley TR. Экогеографические вариации носовых ходов человека. Am J Phys Anthropol.2009; 138: 11–22. pmid: 18623075
- 42. Нобак М.Л., Харвати К., Спур Ф. Климатические изменения носовой полости человека. Am J Phys Anthropol. 2011; 145: 599–614. pmid: 21660932
- 43.
Maddux SD, Yokley TR, Svoma BM, Franciscus RG. Абсолютная влажность и человеческий нос: повторный анализ климатических зон и их влияния на форму и функции носа. Am J Phys Anthropol. 2016; 161: 309–320. pmid: 27374937
- 44. Кек Т., Линдеманн Дж. Численное моделирование и кондиционирование воздуха в носу.GMS Curr Top Otorhinolaryngol Head Neck Surg. 2010; 9: Doc08. pmid: 22073112
- 45. Интхавонг К., Тиан Ц.Ф., Вт JY. Моделирование CFD на способность нагревания в полости носа человека. Ринология. 2007; 842–847.
- 46. Ирмак М.К., Коркмаз А., Эрогул О. Селективное охлаждение мозга, по-видимому, является механизмом, ведущим к черепно-лицевому разнообразию человека, наблюдаемому в разных географических регионах. Мед-гипотезы. 2004; 63: 974–979. pmid: 15504564
- 47. Мэлони СК, Митчелл Д., Блейч Д.Вклад вариабельности каротидного ретата в вариабельность температуры мозга у овец в термонейтральной среде. Am J Physiol Regul Integr Comp Physiol. 2007; 292: R1298–305. pmid: 17082355
- 48.
Митчелл Д., Мэлони С.К., Джессен С., Лабурн Х.П., Камерман П.Р., Митчелл Дж. И др. Адаптивная гетеротермия и избирательное охлаждение мозга у млекопитающих засушливой зоны. Comp Biochem Physiol Часть B Biochem Mol Biol. 2002; 131: 571–585.
- 49. Hetem RS, Strauss WM, Fick LG, Maloney SK, Meyer LCR, Fuller A и др.Избирательное охлаждение мозга арабского орикса (Oryx leucoryx): физиологический механизм борьбы с засушливостью? J Exp Biol. 2012; 215: 3917–24. pmid: 22899527
- 50. Blix AS, Walløe L, Folkow LP. Регуляция температуры головного мозга зимних оленей в условиях теплового стресса. J Exp Biol. 2011; 214: 3850–6. pmid: 22031750
- 51. Crandall CG, Brothers RM, Zhang R, Brengelmann GL, Covaciu L, Jay O, et al. Комментарии по пункту: контрапункт: люди демонстрируют / не демонстрируют избирательное охлаждение мозга во время гипертермии.J Appl Physiol. Американское физиологическое общество; 2011; 110: 575–80. pmid: 21304015
- 52.
Кабанак М. Избирательное охлаждение мозга у человека: «фантазия» или факт? FASEB J. 1993; 7: 1143-1146-1147.
- 53. Ставит Д.А. Красавица и чудовище: механизмы полового отбора у человека. Evol Hum Behav. Эльзевир; 2010. 31: 157–175.
- 54. van Doorn GS, Edelaar P, Weissing FJ. О происхождении видов путем естественного и полового отбора. Наука. 2009; 326: 1704–1707.pmid: 19965377
- 55. Bigham AW, Mao X, Mei R, Brutsaert T, Wilson MJ, Julian CG, et al. Выявление кандидатных локусов положительного отбора для высотной адаптации в андских популяциях. Hum Genomics. 2009; 4: 79–90. pmid: 20038496
- 56. Ву Т., Ли С., член парламента палаты. Тибетцы на большой высоте. Wilderness Environ Med. 2005; 16: 47–54. pmid: 15813148
- 57.
Патерностер Л., Журов А.И., Тома А.М., Кемп Дж. П., Сент-Пуркэн Б., Тимпсон Нью-Джерси и др. Полногеномное ассоциативное исследование трехмерной морфологии лица идентифицирует вариант PAX3, связанный с положением назиона.Am J Hum Genet. Эльзевир; 2012; 90: 478–85. pmid: 22341974
- 58. Лю Ф., ван дер Лийн Ф., Шурманн К., Чжу Г., Чакраварти М.М., Хизи П.Г. и др. Полногеномное ассоциативное исследование выявило пять локусов, влияющих на морфологию лица у европейцев. Гибсон Дж., Редактор. PLoS Genet. Публичная научная библиотека; 2012; 8: e1002932. pmid: 23028347
- 59. Shaffer JR, Orlova E, Lee MK, Leslie EJ, Raffensperger ZD, Heike CL и др. Полногеномное исследование ассоциации выявило множественные локусы, влияющие на нормальную морфологию лица человека.Барш Г.С., редактор. PLOS Genet. Публичная научная библиотека; 2016; 12: e1006149. pmid: 27560520
- 60. Адхикари К., Фуэнтес-Гуахардо М., Квинто-Санчес М., Мендоса-Ревилья Дж., Камило Чакон-Дуке Дж., Акуна-Алонсо В. и др. Полногеномное сканирование ассоциаций вовлекает DCHS2, RUNX2, GLI3, PAX1 и EDAR в изменение лица человека. Nat Commun. Исследования природы; 2016; 7: 11616.
- 61.
Beleza S, Campos J, Lopes J, Araújo II, Hoppfer Almada A, Correia e Silva A и др. Структура примесей и генетическая изменчивость архипелага Кабо-Верде и ее значение для картографических исследований примесей.PLoS One. 2012; 7.
- 62. Нить «TOP / BOT» и аллель «A / B» компании Illumina Inc. — Руководство по методу Illumina для определения цепи и аллеля для анализов GoldenGate и Infinium. (Техническое примечание) [Интернет]. 2006. Доступно: http://www.illumina.com/documents/products/technotes/technote_topbot.pdf
- 63. Перселл С., Нил Б., Тодд-Браун К., Томас Л., Феррейра МАР, Бендер Д. и др. PLINK: набор инструментов для полногеномного анализа ассоциаций и популяционного анализа сцепления.Am J Hum Genet. 2007. 81: 559–75. pmid: 17701901
- 64. Чанг СС, Чоу СС, Теллиер Л.К., Ваттикути С., Перселл С.М., Ли Дж.Дж. PLINK второго поколения: ответ на вызов более крупных и богатых наборов данных. Gigascience. БиоМед Централ Лтд; 2015; 4: 7. pmid: 25722852
- 65.
Candille SI, Absher DM, Beleza S, Bauchet M, McEvoy B, Garrison NA и др. Полногеномные ассоциативные исследования количественно измеренной пигментации кожи, волос и глаз в четырех европейских популяциях.PLoS One. Публичная научная библиотека; 2012; 7: e48294. pmid: 23118974
- 66. Storz JF. Контрастные модели дивергенции количественных признаков и нейтральных ДНК-маркеров: анализ клинальной изменчивости. Mol Ecol. 2002; 11: 2537–2551. pmid: 12453238
- 67. Хе Й, Ли Р., Ван Дж, Бланше С., Лек С. Морфологические изменения среди диких популяций китайского редкого гольяна (Gobiocypris rarus): расшифровка роли эволюционных процессов. Zoolog Sci. 2013; 30: 475–83.pmid: 23721472
- 68. Weir BS, Cockerham CC. Оценка F-статистики для анализа структуры населения. Эволюция (N Y). 1984; 38: 1358–1370.
- 69. Браунинг СР, Браунинг БЛ. Структура популяции может привести к завышению оценок наследуемости на основе SNP. Am J Hum Genet. 2011; 89: 191–193. pmid: 21763486
- 70.
Годдард М. Э., Ли С.Х., Ян Дж., Рэй Н.Р., Вишер П.М. Реакция на потемнение и потемнение. Am J Hum Genet. 2011; 89: 193–195.
- 71.Баран Й., Пасанюк Б., Санкарараман С., Торгерсон Д.Г., Жинью С., Энг С. и др. Быстрый и точный вывод о местной родословной в латиноамериканских популяциях. Биоинформатика. 2012; 28: 1359–1367. pmid: 22495753
- 72. Гальдер I, Шрайвер, доктор медицины. Измерение и использование примесей для изучения генетики сложных заболеваний. Hum Genomics. БиоМед Централ Лтд; 2003; 1: 52. pmid: 15601533
- 73. Кале Д., Уикхэм Х. ggmap: Пространственная визуализация с помощью. Р. Дж. 2013; 5: 144–161.Доступно: http://journal.r-project.org/archive/2013-1/kahle-wickham.pdf
- 74. R Core Team. R: Язык и среда для статистических вычислений. R Фонд статистических вычислений, Вена, Австрия. 2013.
- 75.
Харрис I, Джонс PD, Осборн Т.Дж., Листер DH. Обновленные сетки ежемесячных климатических наблюдений с высоким разрешением — набор данных CRU TS3. 10. Int J Climatol. John Wiley & Sons, Ltd; 2014; 34: 623–642.
- 76. Научная группа ТОМС (1996).TOMS Earth Probe Ультрафиолетовая отражательная способность Ежемесячно L3 Global 1 град x 1,25 градуса широта / долгота Сетка V008, версия 008, Гринбелт, Мэриленд, Центр данных и информационных услуг Годдарда по наукам о Земле (GES DISC) [Интернет]. Доступно: http://disc.sci.gsfc.nasa.gov/datacollection/TOMSEPL3mref_008.html
- 77. Мична П., Вудс М. RNetCDF — Пакет для чтения и записи наборов данных NetCDF. Р. Дж. 2013; 5: 29–35.
- 78. Хиджманс Р. Дж., Эттен Дж. Ван. растр: географический анализ и моделирование с растровыми данными.Пакет R версии 2.0–12. [Интернет]. 2012. Доступно: http://cran.r-project.org/package=raster
- 79. Руссе Ф., Ферди Ж. Б.. Тестирование экологических и генетических эффектов при наличии пространственной автокорреляции. Экография (Мент). 2014; 37: 781–790.
- 80.
Клас П. , Рейджнирс Дж., Шрайвер М.Д., Снайдерс Дж., Суэтенс П., Ниландт Дж. И др. Исследование симметрии соответствия в ушных раковинах человека с возможными последствиями для трехмерного распознавания уха и локализации звука.J Anat. 2015; 226: 60–72. pmid: 25382291
- 81. Пинейро Дж., Бейтс Д., Деброй С., Саркар Д., Команда TRC. nlme: линейные и нелинейные модели смешанных эффектов. Октябрь. 2013. С. 1–3.
 Введение в сравнительную анатомию носа и придаточных пазух носа. Ann R Coll Surg Engl. 1954; 15: 141–171. pmid: 13198060
Введение в сравнительную анатомию носа и придаточных пазух носа. Ann R Coll Surg Engl. 1954; 15: 141–171. pmid: 13198060 pmid: 14350081
pmid: 14350081 , Макэвой Б. П., Гордон С., Хендерс А. К., Найхольт Д. Р. и др. Распространенные SNP объясняют большую часть наследуемости по росту человека. Нат Жене. Nature Publishing Group, подразделение Macmillan Publishers Limited. Все права защищены.; 2010; 42: 565–9. pmid: 20562875
, Макэвой Б. П., Гордон С., Хендерс А. К., Найхольт Д. Р. и др. Распространенные SNP объясняют большую часть наследуемости по росту человека. Нат Жене. Nature Publishing Group, подразделение Macmillan Publishers Limited. Все права защищены.; 2010; 42: 565–9. pmid: 20562875 А., Парра Э., МакКейг П., Мао Х, Ченг К. и др. Генетические доказательства конвергентной эволюции светлой кожи у европейцев и выходцев из Восточной Азии. Mol Biol Evol. 2007; 24: 710–22. pmid: 17182896
А., Парра Э., МакКейг П., Мао Х, Ченг К. и др. Генетические доказательства конвергентной эволюции светлой кожи у европейцев и выходцев из Восточной Азии. Mol Biol Evol. 2007; 24: 710–22. pmid: 17182896 Справочник по антропометрии. Handb Anthr. 2012; 91–114.
Справочник по антропометрии. Handb Anthr. 2012; 91–114. Контрастные модели формы тела и нейтральное генетическое расхождение в морских и озерных популяциях трехиглой колюшки.J Evol Biol. 2006; 19: 1803–12. pmid: 17040377
Контрастные модели формы тела и нейтральное генетическое расхождение в морских и озерных популяциях трехиглой колюшки.J Evol Biol. 2006; 19: 1803–12. pmid: 17040377 Полногеномное обнаружение и характеристика положительного отбора в человеческих популяциях. Природа. 2007; 449: 913–918. pmid: 17943131
Полногеномное обнаружение и характеристика положительного отбора в человеческих популяциях. Природа. 2007; 449: 913–918. pmid: 17943131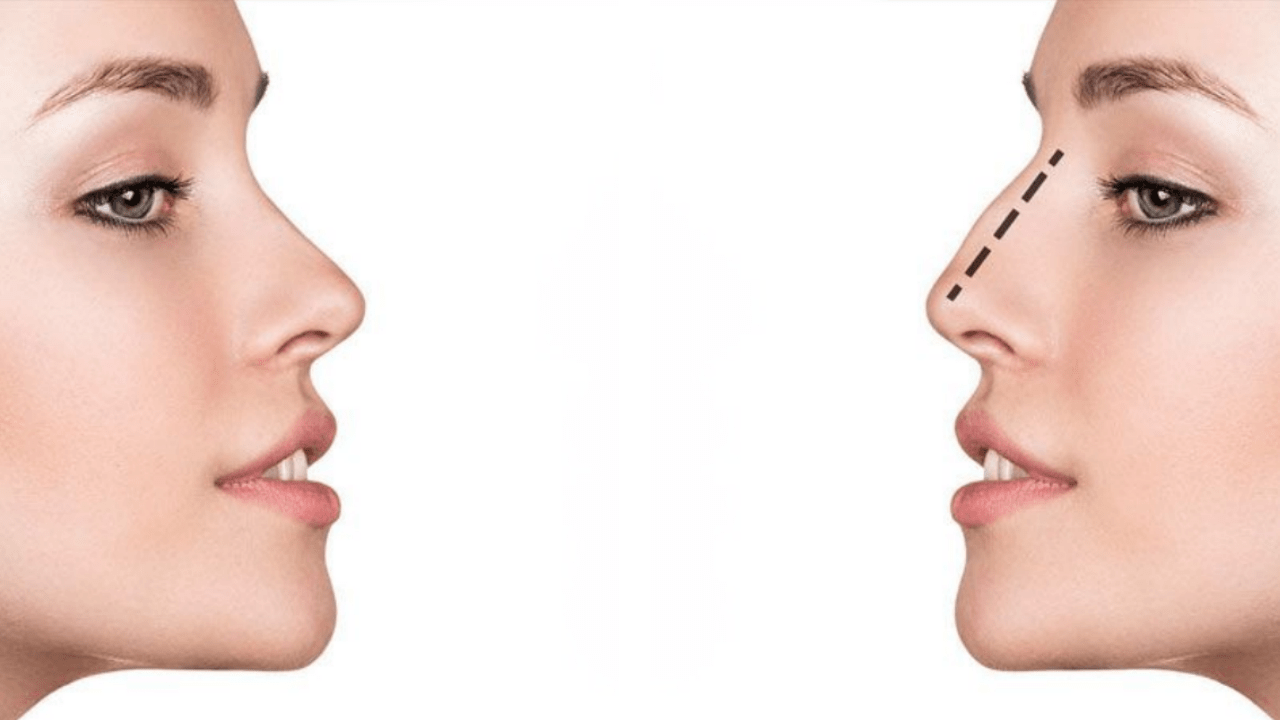 Am J Phys Anthropol. 2016; 161: 309–320. pmid: 27374937
Am J Phys Anthropol. 2016; 161: 309–320. pmid: 27374937 И др. Адаптивная гетеротермия и избирательное охлаждение мозга у млекопитающих засушливой зоны. Comp Biochem Physiol Часть B Biochem Mol Biol. 2002; 131: 571–585.
И др. Адаптивная гетеротермия и избирательное охлаждение мозга у млекопитающих засушливой зоны. Comp Biochem Physiol Часть B Biochem Mol Biol. 2002; 131: 571–585. 1993; 7: 1143-1146-1147.
1993; 7: 1143-1146-1147. pmid: 22341974
pmid: 22341974 Структура примесей и генетическая изменчивость архипелага Кабо-Верде и ее значение для картографических исследований примесей.PLoS One. 2012; 7.
Структура примесей и генетическая изменчивость архипелага Кабо-Верде и ее значение для картографических исследований примесей.PLoS One. 2012; 7. Полногеномные ассоциативные исследования количественно измеренной пигментации кожи, волос и глаз в четырех европейских популяциях.PLoS One. Публичная научная библиотека; 2012; 7: e48294. pmid: 23118974
Полногеномные ассоциативные исследования количественно измеренной пигментации кожи, волос и глаз в четырех европейских популяциях.PLoS One. Публичная научная библиотека; 2012; 7: e48294. pmid: 23118974 Э., Ли С.Х., Ян Дж., Рэй Н.Р., Вишер П.М. Реакция на потемнение и потемнение. Am J Hum Genet. 2011; 89: 193–195.
Э., Ли С.Х., Ян Дж., Рэй Н.Р., Вишер П.М. Реакция на потемнение и потемнение. Am J Hum Genet. 2011; 89: 193–195. 10. Int J Climatol. John Wiley & Sons, Ltd; 2014; 34: 623–642.
10. Int J Climatol. John Wiley & Sons, Ltd; 2014; 34: 623–642. , Рейджнирс Дж., Шрайвер М.Д., Снайдерс Дж., Суэтенс П., Ниландт Дж. И др. Исследование симметрии соответствия в ушных раковинах человека с возможными последствиями для трехмерного распознавания уха и локализации звука.J Anat. 2015; 226: 60–72. pmid: 25382291
, Рейджнирс Дж., Шрайвер М.Д., Снайдерс Дж., Суэтенс П., Ниландт Дж. И др. Исследование симметрии соответствия в ушных раковинах человека с возможными последствиями для трехмерного распознавания уха и локализации звука.J Anat. 2015; 226: 60–72. pmid: 25382291Исследователи говорят, что климат сформировал человеческий нос | Биология человека
Согласно исследованию, исследующему изменчивость человеческой морды, человеческие носы сформировались под воздействием климата.
Исследователи говорят, что их результаты подтверждают теорию о том, что более широкие ноздри развились у популяций, живущих в теплых и влажных условиях, в то время как у людей, живущих в высоких широтах, таких как северная Европа, развились более узкие ноздри в результате адаптации к холодным и сухим условиям.
«Люди долгое время думали, что разница в форме носа у людей во всем мире могла возникнуть в результате естественного отбора из-за климата», — сказал Арслан Заиди, соавтор исследования из Университета штата Пенсильвания. Но в то время как предыдущие исследования были основаны на измерениях человеческих черепов, по его словам, в новом исследовании изучалась сама форма носа.
В журнале Plos Genetics исследователи из США, Ирландии и Бельгии описывают, как они начали распознавать вариации формы носа, используя трехмерную визуализацию лица для проведения множества измерений у 476 добровольцев из Южной Азии, Восточной Азии и Западной Африки. и североевропейское происхождение.
Результаты показали, что только две из семи черт, связанных с носом, различались между популяциями больше, чем можно было бы ожидать из-за воздействия случайных, случайных изменений генетической структуры с течением времени. Авторы говорят, что это предполагает, что на вариации этих черт повлиял естественный отбор.
После дальнейшего анализа, основанного на данных участников западноафриканского и европейского происхождения, подтверждающего, что форма носа в значительной степени наследуется, команда попыталась выяснить, существует ли связь между формой носа и климатом.
Синие прямоугольники представляют собой аллели более узких носовых ходов, а красные прямоугольники представляют аллели более широких носовых ходов. Цвета между ними представляют промежуточные фенотипы. Цветовая шкала представляет климатические изменения температуры и влажности, чтобы показать, что более узкие ноздри предпочтительны в прохладно-сухом климате, а более широкие — в жарком и влажном климате. Фотография: Арслан А. Заиди и его коллеги Для этого они исследовали размеры носа у 140 женщин западноафриканского, восточноазиатского, североевропейского или южноазиатского происхождения, чьи родители жили в регионе, который соответствовал этой родословной. .Затем были использованы места рождения родителей каждого участника для присвоения каждой женщине значений температуры и влажности, которые приблизительно отражали климат, в котором жили их недавние предки.
Результаты показали, что ширина ноздри связана с температурой и абсолютной влажностью: участники, чьи предки жили в теплом и влажном климате, в среднем имели более широкие ноздри, чем те, чьи предки жили в прохладно-сухом климате.
Это, по словам Заиди, могло быть связано с тем, что более узкие носовые ходы помогают увеличить влажность воздуха и согреть его — бонус для тех, кто живет в высоких широтах.«Холодный и сухой воздух вреден для наших внутренних дыхательных путей», — сказал Заиди. Но, добавляет он, «не существует универсально« хорошей »или« лучшей »формы — ваши предки были адаптированы к окружающей среде, в которой они жили». Заиди признает, что исследование не исключает возможности того, что другие факторы, помимо климата, могут иметь место. в игре, не в последнюю очередь то, что разные группы населения могут считать определенные черты более или менее привлекательными, а это означает, что за различиями может стоять половой отбор.
Заиди считает, что устранение человеческих приспособлений очень важно для понимания риска заболеваний. «По мере того, как мы становимся все более глобальным сообществом, по мере того как мы перемещаемся по миру, мы будем сталкиваться с климатом, к которому мы не адаптированы», — сказал он, предполагая, что переезд в совершенно другой климат может увеличить риск респираторных заболеваний. Но, добавил он, «это не обязательно может быть правдой по разным причинам, например, из-за современной медицины и того факта, что наш нынешний климат сильно отличается от того, что было раньше».
«По мере того, как мы становимся все более глобальным сообществом, по мере того как мы перемещаемся по миру, мы будем сталкиваться с климатом, к которому мы не адаптированы», — сказал он, предполагая, что переезд в совершенно другой климат может увеличить риск респираторных заболеваний. Но, добавил он, «это не обязательно может быть правдой по разным причинам, например, из-за современной медицины и того факта, что наш нынешний климат сильно отличается от того, что было раньше».
Норин фон Крамон-Таубадель из Университета Буффало, штат Нью-Йорк, которая не принимала участия в исследовании, сказала, что климатическое давление вряд ли будет полной историей, когда дело доходит до возможной адаптации носа.
«Хотя ширина носа может быть предметом естественного отбора, неясно, обусловлено ли это климатом», — говорит она, указывая на то, что связь исследования между климатом и формой носа исчезает, когда северные европейцы удаляются из анализа.
Более того, говорит фон Крамон-Таубадель, исследование сосредоточено только на внешних характеристиках носа, хотя его внутренняя структура также важна, когда дело касается кондиционирования воздуха. «Таким образом, хотя это исследование проливает свет на эволюцию внешней формы носа, оно не обязательно затрагивает аспекты анатомии носа, которые являются наиболее важными для понимания климатической адаптации», — сказала она.
«Таким образом, хотя это исследование проливает свет на эволюцию внешней формы носа, оно не обязательно затрагивает аспекты анатомии носа, которые являются наиболее важными для понимания климатической адаптации», — сказала она.
Заиди сказал, что необходимы дальнейшие исследования для проверки связи между климатом и формой носа, включая проведение аналогичных исследований с участниками из более широкого круга предков, включая коренных американцев и инуитов, и выявление генов, лежащих в основе различий в форме носа.
Но, добавляет он, текущее исследование подчеркивает важный момент.
«Между людьми из разных популяций больше сходства, чем различий», — сказал Заиди.«Это то, что мы обнаруживаем и в нашей статье — на самом деле большинство черт не сильно различаются между популяциями».
Как климат помогает объяснить, как ваш нос приобрел форму
Новое исследование меняет наше понимание того, как влажность повлияла на эволюцию человеческого носа.
(Наука изнутри) — Хотя наши носы удобны для того, чтобы уловить разницу между, скажем, цветущим трупным цветком и кедровым деревом, они также играют часто упускаемую из виду роль: кондиционер.
Когда мы вдыхаем, наши носы нагревают и кондиционируют воздух, подготавливая его к обмену кислорода и углекислого газа в легких. Если воздух слишком сухой, его помогает увлажнять нос. Этот процесс также предотвращает высыхание респираторных тканей, что может предотвратить развитие инфекций.
Предыдущее исследование показало, что, поскольку людям необходимо регулировать температуру и влажность воздуха, входящего в их тело, носы в более теплом и прохладном климате приобрели разные формы.Теперь новое исследование уточняет, как климат помогает объяснить, как ваш нос приобрел форму.
Исследование, опубликованное в июне в American Journal of Physical Anthropology , может привести к лучшему пониманию эволюции человеческого лица и того, почему черты лица различаются у разных этнических групп.
«Это важное исследование, в котором излагаются потенциальные климатические переменные, на которых мы должны сосредоточиться», — сказал Натан Холтон, биологический антрополог из Университета Айовы в Айова-Сити, который не участвовал в исследовании.
В прошлых исследованиях взаимосвязи между климатом и формой носа обычно использовались измерения относительной влажности — отношения количества водяного пара в воздухе к количеству пара, которое он действительно может удерживать, — и температуры. В результате они склонны классифицировать население по четырем климатическим категориям: холодно-влажный, холодно-сухой, горячий-влажный и жарко-сухой.
Новое исследование, однако, основано на предыдущем исследовании 1950-х годов и показывает, что абсолютная влажность — мера того, сколько водяного пара на самом деле присутствует в воздухе — имеет большее значение при попытке выяснить, как климат влияет на форму носа.
Чтобы понять, почему это так, рассмотрим зиму в Новой Англии. Хотя в холодный день может быть высокая относительная влажность, эти условия не всегда считаются «влажными». Это потому, что при низких температурах воздух не может удерживать много воды, объяснил Скотт Мэддакс, ведущий автор исследования и биологический антрополог, работающий сейчас в Научном центре здоровья Университета Северного Техаса в Форт-Уэрте.
Итак, если килограмм этого холодного воздуха может вместить не более двух граммов воды и имеет относительную влажность 90 процентов, общее количество пригодной для дыхания влаги в этой части воздуха все равно будет лишь немногим меньше двух граммов, «что — сказал Мэддукс.
Слабая влагоудерживающая способность холодного воздуха означает, что люди, живущие в холодно-сухих и холодно-влажных средах, испытывают одинаковые потребности в нагревании и увлажнении носа. Различение этих двух климатических условий может ввести в заблуждение сыщиков, пытающихся выяснить, почему мы выглядим именно так.
В прошлом антропологи предполагали, что относительно более узкий нос у некоторых европейцев — это приспособление к холодной и сухой среде, в которой жили их предки. Это потому, что более тонкая форма помогает увеличить степень соприкосновения воздуха со слизистой оболочкой и кровеносными сосудами внутри носа, где на самом деле происходит обмен тепла и влаги.
Более того, антропологи обычно думали, что люди, живущие в жарких и влажных средах, подобных тем, что находятся у экватора, не испытывают особого избирательного давления на нос. Это потому, что, как говорится, воздух уже достаточно теплый и влажный, и его не нужно модифицировать. Только по мере того, как вы удаляетесь от экватора, давление, необходимое для кондиционирования воздуха, увеличивается, и нос меняется соответственно.
Это потому, что, как говорится, воздух уже достаточно теплый и влажный, и его не нужно модифицировать. Только по мере того, как вы удаляетесь от экватора, давление, необходимое для кондиционирования воздуха, увеличивается, и нос меняется соответственно.
Но «мы действительно не думаем, что это правда», — сказал Мэддакс.«Мы думаем, что существует некоторое давление, чтобы потенциально изменить нос немного иначе, чем то, что может происходить в холодной среде».
Их результаты подтверждают идею о том, что в жарких и влажных средах, которые снижают способность нашего тела выделять тепло через потоотделение, нос может реагировать на это давление и со временем приобретает более широкую форму, что обеспечивает большую потерю тепла и может помочь сохранить мозг прохладным, сказал он.
Антропологи все еще работают над установлением таких связей, сказал Холтон.
Это отчасти потому, что формы носа обычно классифицируются на основе внешнего скелета и даже мясистой части носа, потому что это легко измерить. Но большая часть нагрева и увлажнения воздуха происходит во внутренней части носа, называемой носовой полостью, и ученые до сих пор не понимают, в какой степени взаимосвязаны внешняя и внутренняя части.
Но большая часть нагрева и увлажнения воздуха происходит во внутренней части носа, называемой носовой полостью, и ученые до сих пор не понимают, в какой степени взаимосвязаны внешняя и внутренняя части.
Выявление этой связи — например, путем компьютерной томографии черепов живых людей — является следующим шагом, потому что «именно здесь мы действительно должны ожидать увидеть эти отношения между [формой носа] и климатом», — сказал Мэддакс.
Исследователи проанализировали климатические данные с 1901 по 2013 год, чтобы создать глобальные карты среднегодовой температуры, относительной влажности и абсолютной влажности. Для дальнейшего изучения влияния этих факторов на форму носа они использовали данные, собранные в 1923 году у более чем 15 000 человек со всего мира. (Они отмечают, что предыдущие исследования показали, что недавние климатические условия обеспечивают разумное сравнение климата, с которым сталкивались более ранние современные человеческие популяции.)
Но климат, конечно, не единственный фактор, который может объяснить, почему наши клювы варьируются от орлиных до выпуклых. Исследование, проведенное в журнале Nature Communications в мае прошлого года, выявило пять генов, контролирующих форму носа и подбородка человека. По словам Холтона, критически важно понять лежащую в основе генетику, в том числе выяснить, как условия окружающей среды могли повлиять на эволюцию самих этих генов.
Исследование, проведенное в журнале Nature Communications в мае прошлого года, выявило пять генов, контролирующих форму носа и подбородка человека. По словам Холтона, критически важно понять лежащую в основе генетику, в том числе выяснить, как условия окружающей среды могли повлиять на эволюцию самих этих генов.
Это будет шагом к разгадке тайны, мучившей ученых на протяжении многих лет: как эволюционировало человеческое лицо.
«Большая часть любого лица, не только человеческого лица, — это нос», — сказал Холтон.«Буквально это центральный компонент черепа, поэтому понимание того, что происходит с носом, может многое рассказать нам о том, что происходит с остальной частью лица».
Какая форма и размер носа подходят вам? — Доктор Брент
Ринопластика (часто называемая «пластикой носа») — отличная процедура для всех, кто недоволен размером или формой своего носа. Это одна из наиболее распространенных косметических процедур, поскольку ежегодно ее проходят миллионы людей.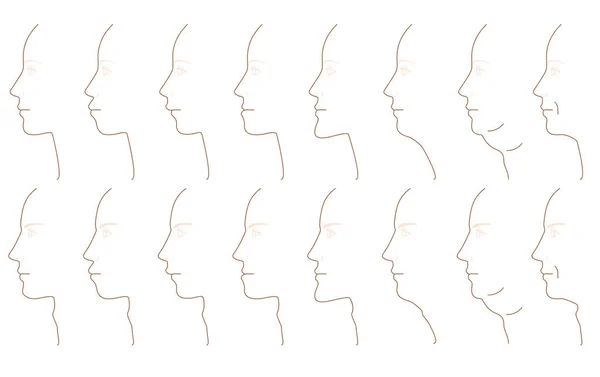 Одна из наиболее сложных частей процедуры — это выбор размера и формы, которые вы хотите, поскольку строение лица каждого человека уникально.Это может быть действительно ошеломляющим, поскольку вы, возможно, захотите поменять нос, но не знаете, что именно вам подойдет лучше всего. Итак, какая форма и размер носа подходят именно вам?
Одна из наиболее сложных частей процедуры — это выбор размера и формы, которые вы хотите, поскольку строение лица каждого человека уникально.Это может быть действительно ошеломляющим, поскольку вы, возможно, захотите поменять нос, но не знаете, что именно вам подойдет лучше всего. Итак, какая форма и размер носа подходят именно вам?
Какая форма и размер носа подходят вам?
Вот почему так важно найти сертифицированного пластического хирурга, который готов проводить с вами время во время консультации. Ниже вы найдете дополнительную информацию о ринопластике, а также о том, какая форма и размер носа вам подходят.
Консультации по ринопластике и рекомендации
Одна из важных особенностей ринопластики — это то, что она может быть косметической, по состоянию здоровья или даже того и другого.Поскольку ринопластика — это операция, которая изменяет форму или размер вашего носа, важно обсудить это с вашим сертифицированным хирургом на консультации. Ваш нос состоит из кожи, костей и хрящей, и каждый из них, вероятно, будет изменен во время процедуры.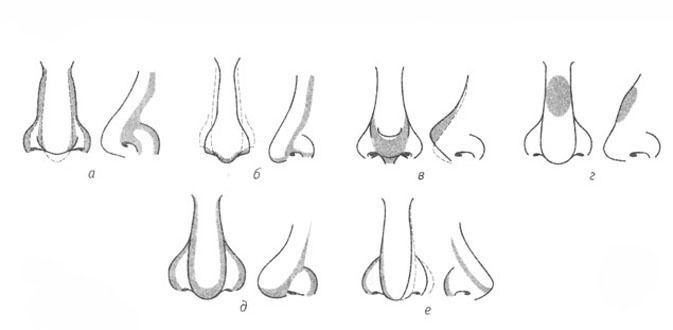 Ниже вы найдете важные вещи, которые нужно учитывать, и что именно обсуждать на консультации.
Ниже вы найдете важные вещи, которые нужно учитывать, и что именно обсуждать на консультации.
Верхний нос:
Общие изменения, которые происходят в верхней части носа, часто связаны с уменьшением выпуклости или сужением более широкого посаженного носа.Если вы хотите выпрямить или сузить нос, рекомендуется заняться верхней частью носа. Что касается пропорций, вы не хотите, чтобы верхняя часть носа была больше нижней.
Средний нос:
Для тех, у кого более широкий нос, эту область можно проредить. Однако важно не худеть слишком сильно, так как это может повлиять на ваше дыхание. Как и в случае с верхней частью носа, рекомендуется проредить верхнюю и среднюю части носа, поскольку вы не хотите, чтобы они были больше, чем нижняя часть носа.
Кончик носа:
Если кончик вашего носа отвисает или высовывается наружу, вы можете уменьшить эту область. Однако очень важно получить опытного хирурга, так как это сложная часть процедуры. Часто сама процедура, помимо процесса заживления, определяет успех лечения этой области вашего носа.
Назальные дыхательные пути:
Из-за важности носового дыхания процедура может быть полезной для здоровья. Если у вас всегда были проблемы с дыханием через нос или у вас была аллергия, поговорите со своим хирургом на консультации о своих ожиданиях.
Процедура ринопластики и восстановление
После консультации вы сможете записаться на прием по поводу пластики носа в Беверли-Хиллз. В день процедуры вам сделают анестезию. Затем ваш хирург сделает серию надрезов для достижения желаемого эстетического результата, удалив определенные части вашего носа в дополнение к его изменению. В некоторых случаях в нос может быть добавлен хрящ. После того, как изменения будут внесены, хирург стратегически поместит вашу кожу поверх недавно измененного носа.Процедура занимает от одного до двух часов в зависимости от сложности ваших конкретных потребностей.
После того, как вы выйдете из операции, ваш хирург наложит шину на ваш нос, чтобы помочь ему сохранить свою новую форму во время заживления. У вас также будут носовые мешки или шины внутри носа, чтобы он не свалился. В течение нескольких дней после операции вы можете испытывать боль, синяки, кровотечение или дренаж. Это нормально и должно пройти через короткое время. Очень важно в течение первых недель или двух сосредоточиться на отдыхе, чтобы не только ваш нос, но и все ваше тело восстановились после операции.
У вас также будут носовые мешки или шины внутри носа, чтобы он не свалился. В течение нескольких дней после операции вы можете испытывать боль, синяки, кровотечение или дренаж. Это нормально и должно пройти через короткое время. Очень важно в течение первых недель или двух сосредоточиться на отдыхе, чтобы не только ваш нос, но и все ваше тело восстановились после операции.
Лучший хирург в Беверли-Хиллз
Если вы недовольны видом или формой своего носа, свяжитесь с доктором Брентом и его командой. Доктор Брент Моэллекен — известный косметический хирург с многолетним опытом в пластической и реконструктивной хирургии, включая ринопластику. Он является одним из самых высококвалифицированных хирургов в стране и прошел обучение в Гарварде, Йельском университете и Калифорнийском университете в Лос-Анджелесе. Он также был включен в список 10 лучших пластических хирургов Америки по версии журнала Vogue. Свяжитесь с ведущим косметическим хирургом Беверли-Хиллз сегодня для консультации!
Расследование случая формы человеческого носа и адаптации к климату
Abstract
Эволюционные причины вариаций формы носа в разных популяциях людей являются предметом постоянных дискуссий. Важной функцией носа и носовой полости является кондиционирование вдыхаемого воздуха до того, как он достигнет нижних дыхательных путей. По этой причине считается, что наблюдаемые различия в форме носа среди популяций не просто результат генетического дрейфа, но могут быть адаптацией к климату. Чтобы ответить на вопрос, является ли местная адаптация к климату ответственной за дивергенцию формы носа у популяций, мы используем сравнения Qst – Fst, чтобы показать, что ширина ноздрей и ширина крылового основания более дифференцированы в популяциях, чем ожидалось при одном только генетическом дрейфе.Чтобы проверить, связана ли эта дифференциация с адаптацией к климату, мы сравнили пространственное распределение этих переменных с глобальным распределением температуры, абсолютной влажности и относительной влажности. Мы обнаружили, что ширина ноздрей коррелирует с температурой и абсолютной влажностью, но не с относительной влажностью. Мы пришли к выводу, что некоторые аспекты формы носа действительно могли быть обусловлены местной адаптацией к климату. Однако мы думаем, что это упрощенное объяснение очень сложной эволюционной истории, которая, возможно, также включала другие ненейтральные силы, такие как половой отбор.
Важной функцией носа и носовой полости является кондиционирование вдыхаемого воздуха до того, как он достигнет нижних дыхательных путей. По этой причине считается, что наблюдаемые различия в форме носа среди популяций не просто результат генетического дрейфа, но могут быть адаптацией к климату. Чтобы ответить на вопрос, является ли местная адаптация к климату ответственной за дивергенцию формы носа у популяций, мы используем сравнения Qst – Fst, чтобы показать, что ширина ноздрей и ширина крылового основания более дифференцированы в популяциях, чем ожидалось при одном только генетическом дрейфе.Чтобы проверить, связана ли эта дифференциация с адаптацией к климату, мы сравнили пространственное распределение этих переменных с глобальным распределением температуры, абсолютной влажности и относительной влажности. Мы обнаружили, что ширина ноздрей коррелирует с температурой и абсолютной влажностью, но не с относительной влажностью. Мы пришли к выводу, что некоторые аспекты формы носа действительно могли быть обусловлены местной адаптацией к климату. Однако мы думаем, что это упрощенное объяснение очень сложной эволюционной истории, которая, возможно, также включала другие ненейтральные силы, такие как половой отбор.
Авторское резюме
Изучение адаптации человека имеет важное значение для нашего понимания этиологии болезни. Эволюционные исследования того, почему определенные фенотипы заболеваний, такие как серповидноклеточная анемия и непереносимость лактозы, возникают с разной скоростью в разных популяциях, привели к лучшему пониманию вовлеченных генетических и экологических факторов риска. Точно так же исследования географического распределения пигментации кожи продолжают давать важные подсказки относительно риска дефицита витамина D и рака кожи.Здесь мы исследуем, было ли изменение формы носа у разных популяций вызвано региональными различиями в климате. Мы обнаружили, что вариации как в ширине ноздрей, так и в ширине крылового основания, по-видимому, ускоренно разошлись среди человеческих популяций. Мы также обнаружили, что геопространственное распределение ширины ноздрей коррелирует с температурой и абсолютной влажностью, но не с относительной влажностью. Наши результаты подтверждают утверждение о том, что местная адаптация к климату могла сыграть роль в эволюции различий формы носа в разных популяциях людей.
Наши результаты подтверждают утверждение о том, что местная адаптация к климату могла сыграть роль в эволюции различий формы носа в разных популяциях людей.
Введение
Форма носа, как и многих других частей лица, варьируется как внутри, так и среди людей. Например, расстояние между носовыми крыльями (крыльями носа) значительно больше у людей западноафриканского, южноазиатского и восточноазиатского происхождения по сравнению с людьми европейского происхождения [1]. Носовой указатель (ширина / высота носового отверстия черепа) также, как известно, значительно различается в разных популяциях [1,2]. Неясно, вызваны ли эти популяционные различия в форме носа в первую очередь генетическим дрейфом или естественным отбором.
Жизненно важная функция носа — нагревать вдыхаемый воздух до температуры тела и насыщать его водяным паром, прежде чем он достигнет нижних дыхательных путей [3]. Фактически, вдыхаемый воздух достигает 90% от требуемых уровней температуры и влажности еще до того, как достигает носоглотки, что делает полость носа главным кондиционирующим аппаратом в дыхательных путях [4,5]. Это кондиционирование поддерживает надлежащее функционирование мукоцилиарного аппарата, который улавливает частицы и патогены и удаляет их из дыхательных путей.Низкая влажность в дыхательных путях приводит к нарушению мукоцилиарной функции и увеличивает риск инфекций как верхних, так и нижних дыхательных путей [6]. Большая часть кондиционирования воздуха происходит, когда он проходит через носовые раковины, стенки которых выстланы кровеносными сосудами и слизью, производящими бокаловидные клетки [4]. Исследования показали, что эффективность процесса кондиционирования зависит от динамики потока вдыхаемого воздуха, которая, в свою очередь, зависит от геометрии носовой полости и входных отверстий [5,7].Поскольку нос выполняет функцию кондиционера, предполагается, что различия в форме носа у разных групп населения могли быть вызваны местной адаптацией к климату [2,3,8].
Это кондиционирование поддерживает надлежащее функционирование мукоцилиарного аппарата, который улавливает частицы и патогены и удаляет их из дыхательных путей.Низкая влажность в дыхательных путях приводит к нарушению мукоцилиарной функции и увеличивает риск инфекций как верхних, так и нижних дыхательных путей [6]. Большая часть кондиционирования воздуха происходит, когда он проходит через носовые раковины, стенки которых выстланы кровеносными сосудами и слизью, производящими бокаловидные клетки [4]. Исследования показали, что эффективность процесса кондиционирования зависит от динамики потока вдыхаемого воздуха, которая, в свою очередь, зависит от геометрии носовой полости и входных отверстий [5,7].Поскольку нос выполняет функцию кондиционера, предполагается, что различия в форме носа у разных групп населения могли быть вызваны местной адаптацией к климату [2,3,8].
Есть несколько проблем при проверке этой гипотезы. Мы знаем, что форма носа существенно различается среди человеческих популяций, как по внешней морфологии носа, так и по основной морфологии черепа [1,2,9]. Хотя это можно объяснить адаптацией к местному давлению отбора, это также можно объяснить тем фактом, что фенотипические различия между географически удаленными популяциями могут возникать просто из-за генетического дрейфа.Таким образом, чтобы использовать дивергентный отбор в качестве объяснения, необходимо продемонстрировать, что наблюдаемые вариации формы носа в разных популяциях людей больше, чем ожидаемые при одном генетическом дрейфе. Это можно сделать с помощью статистики Qst, которая измеряет степень генетической дифференциации, лежащей в основе количественного признака [10]. В принципе, ожидается, что Qst нейтрально эволюционирующего признака будет соответствовать распределению Fst нейтрально развивающихся локусов [11]. Таким образом, когда Qst намного больше, чем Fst, расхождение признаков превышает нейтральные ожидания и может быть отнесено на счет дивергентного отбора [11].Проблема с Qst заключается в том, что его расчет требует знания аддитивных генетических вариаций внутри и между популяциями.
Хотя это можно объяснить адаптацией к местному давлению отбора, это также можно объяснить тем фактом, что фенотипические различия между географически удаленными популяциями могут возникать просто из-за генетического дрейфа.Таким образом, чтобы использовать дивергентный отбор в качестве объяснения, необходимо продемонстрировать, что наблюдаемые вариации формы носа в разных популяциях людей больше, чем ожидаемые при одном генетическом дрейфе. Это можно сделать с помощью статистики Qst, которая измеряет степень генетической дифференциации, лежащей в основе количественного признака [10]. В принципе, ожидается, что Qst нейтрально эволюционирующего признака будет соответствовать распределению Fst нейтрально развивающихся локусов [11]. Таким образом, когда Qst намного больше, чем Fst, расхождение признаков превышает нейтральные ожидания и может быть отнесено на счет дивергентного отбора [11].Проблема с Qst заключается в том, что его расчет требует знания аддитивных генетических вариаций внутри и между популяциями. Их можно надежно оценить только с помощью экспериментов «обычного сада», в которых влияние окружающей среды на фенотип можно эффективно контролировать. Поскольку такие эксперименты невозможны на людях, вывод на основе Qst относительно дивергентного отбора по фенотипам человека требует принятия реалистичных предположений о наследуемости рассматриваемых фенотипов.
Их можно надежно оценить только с помощью экспериментов «обычного сада», в которых влияние окружающей среды на фенотип можно эффективно контролировать. Поскольку такие эксперименты невозможны на людях, вывод на основе Qst относительно дивергентного отбора по фенотипам человека требует принятия реалистичных предположений о наследуемости рассматриваемых фенотипов.
В нескольких исследованиях использовался этот подход и было обнаружено, что, хотя большинство аспектов черепа, кажется, эволюционируют нейтрально, форма носового отверстия, по-видимому, более дифференцирована в разных популяциях людей, чем ожидалось при генетическом дрейфе [9,12,13].Совсем недавно также сообщалось, что расхождение в форме внешнего носа, по крайней мере, между европейцами и китайцами-ханьцами, также превышает нейтральные ожидания [14]. Хотя это может быть правдой, в большинстве этих исследований использовались антиконсервативные предположения о наследственности, которые переоценивают генетическую дифференциацию, лежащую в основе признака, и, таким образом, приводят к неверным выводам относительно относительной роли отбора и дрейфа в движущих фенотипических различиях между популяциями.
Здесь мы провели исследование вариаций формы носа в свете количественной генетической теории, чтобы выяснить, превышает ли вариация формы носа среди популяций нейтральные ожидания.При этом мы обсуждаем методологические проблемы, связанные с решением таких вопросов у людей, и показываем, как ограничения предыдущих исследований могут быть устранены с помощью последних достижений в статистической генетике. В качестве принципиального доказательства мы сравниваем дифференциацию формы носа с двумя сильно наследуемыми морфологическими признаками, которые, как известно, имеют полигенную основу; а именно рост и пигментация кожи [15–18]. Как рост, так и пигментация кожи существенно различаются внутри и между популяциями людей, и, как полагают, в различных популяциях они подвергаются положительному отбору [19–22].Пигментация кожи также является хорошим примером фенотипа, который, как известно, развился в ответ на меняющееся в географическом пространстве давление отбора: воздействие ультрафиолетового излучения B (UVB) на уровне земли [23].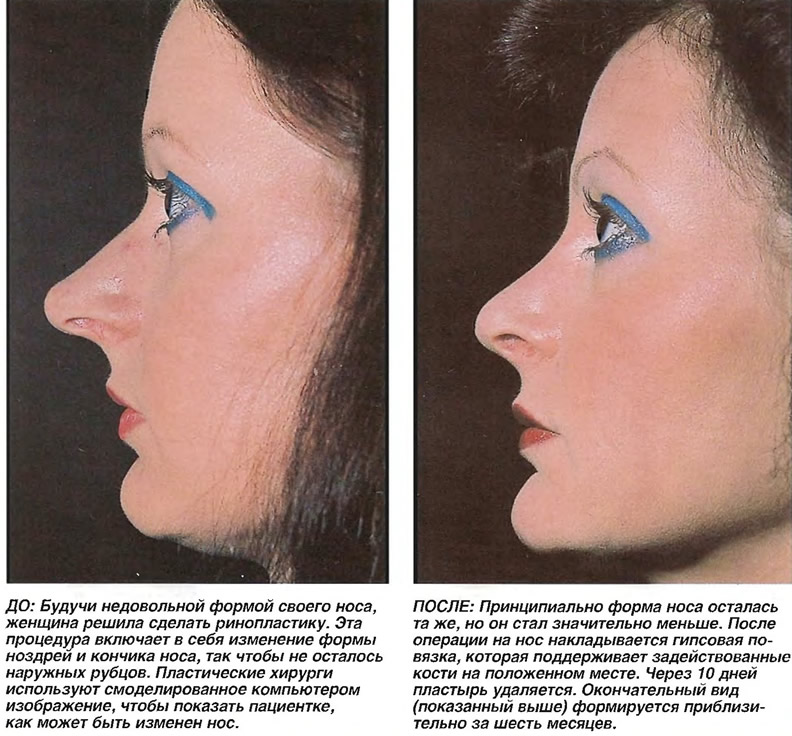 Наконец, мы проверяем, зависит ли клинальная вариация в аспектах формы носа, которая, по-видимому, ускоряется в разных популяциях, от географических вариаций температуры и влажности, чтобы определить, вызвано ли это расхождение давлением климатического отбора.
Наконец, мы проверяем, зависит ли клинальная вариация в аспектах формы носа, которая, по-видимому, ускоряется в разных популяциях, от географических вариаций температуры и влажности, чтобы определить, вызвано ли это расхождение давлением климатического отбора.
Результаты
Описание вариации формы носа
Для количественной оценки вариации формы носа мы сначала сделали трехмерные изображения лиц участников с высоким разрешением, используя систему 3dMD Face ( 3dMD Atlanta , GA ).Были размещены пять ориентиров (два на внутреннем углу глаз, два на внешних углах рта и один на кончике носа), чтобы установить ориентацию лица. Пространственно плотная сетка из 7 150 квази-ориентиров (QL) была нанесена на каждое изображение и его отражение. Обобщенное наложение Прокруста [24] было выполнено на обоих наборах изображений (исходном и отраженном) для устранения различий в положении и ориентации. Координаты Прокруста исходного и отраженного изображений для каждого участника затем были усреднены, чтобы удалить эффекты двусторонней асимметрии, как указано в Claes et al . (2014) [25]. Область носа, которая состоит из 709 из 7 150 QL, была выбрана для последующих анализов.
(2014) [25]. Область носа, которая состоит из 709 из 7 150 QL, была выбрана для последующих анализов.
Мы использовали линейные расстояния и площади, чтобы охарактеризовать форму носа (). Линейные расстояния (измеренные в мм) были рассчитаны с использованием семи стандартных антропометрических ориентиров: i. назион (п), II. пронасале (прн), iii. subnasale (sn), iv. Кривизна левой крыльчатки (al -1 ), v. Искривление правой крыльчатки (al r ), vi. левое основание сигнализатора (ac l ) и vii.Правое основание крыльев (ac r ) () [26,27]. Эти семь ориентиров были размещены на пространственно-плотной сетке из 709 QL, что облегчило автоматическое размещение на каждой грани. Мы использовали координаты x, y и z этих ориентиров для вычисления пяти евклидовых расстояний: i. ширина ноздрей (al l −al r ), ii. ширина основания alar (ac l −ac r ), iii. высота носа (n – sn), iv. длина носового гребня (n – prn) и v. выступ кончика носа (sn – prn) (). Мы также вычислили две площади (измеренные в мм 2 ): i.общая внешняя площадь носа и ii. средняя площадь ноздри () (Методы).
выступ кончика носа (sn – prn) (). Мы также вычислили две площади (измеренные в мм 2 ): i.общая внешняя площадь носа и ii. средняя площадь ноздри () (Методы).
A) Места ориентиров, которые использовались для расчета линейных расстояний, показаны красными точками. Б) Для описания формы носа использовались пять линейных расстояний (красные линии) и две площади поверхности (красная сетка). Линейные расстояния рассчитывались по трехмерным координатам ориентиров (красные точки). Площадь поверхности рассчитывалась как сумма многоугольников, выделенных красным.
показывает распределение аспектов формы носа у мужчин и женщин из четырех групп населения: Западной Африки, Восточной Азии, Южной Азии и Северной Европы. Эти группы были определены на основе генетического и самооценочного происхождения (методы). также показано распределение роста и индекс меланина, показатель уровня пигментации кожи, полученный с помощью спектрофотометрии отражательной способности (измеренный на внутренней стороне верхней части рук; см. Методы) для сравнения. Размер выборки, среднее значение и стандартное отклонение для каждого фенотипа представлены в.Одно ясное наблюдение заключается в том, что все аспекты формы носа, по крайней мере, рассматриваемые здесь, в значительной степени сексуально диморфны (,). У мужчин в среднем более широкие ноздри, более длинные носовые гребни, более выступающие наружу кончики носа, более крупные ноздри и большая общая площадь внешней поверхности по сравнению с женщинами (,). Все размеры носа также значительно различаются в разных популяциях (таблица S1). Исходя из этого, отметим некоторые общие закономерности. Например, ширина ноздрей и ширина аларного основания самые большие в W.Африканцы и самые мелкие на Севере европейцы. Эти результаты согласуются с предыдущими наблюдениями [1,2]. Выступ кончика носа больше всего у североевропейцев и меньше всего у западноафриканцев и восточноазиатцев. E. Азиаты также имеют самые маленькие носы с точки зрения площади внешней поверхности.
Методы) для сравнения. Размер выборки, среднее значение и стандартное отклонение для каждого фенотипа представлены в.Одно ясное наблюдение заключается в том, что все аспекты формы носа, по крайней мере, рассматриваемые здесь, в значительной степени сексуально диморфны (,). У мужчин в среднем более широкие ноздри, более длинные носовые гребни, более выступающие наружу кончики носа, более крупные ноздри и большая общая площадь внешней поверхности по сравнению с женщинами (,). Все размеры носа также значительно различаются в разных популяциях (таблица S1). Исходя из этого, отметим некоторые общие закономерности. Например, ширина ноздрей и ширина аларного основания самые большие в W.Африканцы и самые мелкие на Севере европейцы. Эти результаты согласуются с предыдущими наблюдениями [1,2]. Выступ кончика носа больше всего у североевропейцев и меньше всего у западноафриканцев и восточноазиатцев. E. Азиаты также имеют самые маленькие носы с точки зрения площади внешней поверхности.

Рост измеряется в сантиметрах, а индекс меланина измеряется в процентах отражательной способности (методы). Линейные расстояния измеряются в миллиметрах (мм), а площадь измеряется в мм. 2 .Точки представляют собой индивидуальные наблюдения, а цвет прямоугольных диаграмм и точек представляет пол, причем синий цвет указывает на мужчин, а красный — на женщин.
Таблица 1
Размер выборки, среднее и стандартное отклонение фенотипов по полу и популяции.
| Население | Пол | N a | Высота (см) | Индекс меланина (% отражения) | Ширина ноздрей (мм) | Ширина основания носа (мм) | Высота носа (мм) | мм) | Длина носового гребня (мм) | Выступ кончика носа (мм) | Площадь внешней поверхности (мм) | Площадь ноздри (мм) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
W. Африканец Африканец | Мужской | 10, 6, 10 | 179,89 b (6,48) | 59,23 (9,68) | 45,36 (2,74) | 45,31 (2,61) | 51,31 (2,36) | 469 | 15,94 (1,44) | 1782,13 (122,92) | 70,17 (10,41) | |
| W. Африканский | Женский | 30, 28, 30 | 164,68 (8,61) | 60,27 (2,95) | 40,57 (2,56) | 49. 15 (3,04) 15 (3,04) | 43,82 (2,76) | 14,43 (0,98) | 1477,42 (167,39) | 57,18 (9,75) | ||
| E. Азиатский | Мужской | 43, 33, 43 | 43, 33, 43 | )31,42 (2,94) | 38,55 (2,28) | 39,98 (2,31) | 52,35 (2,52) | 47,14 (2,59) | 15,81 (1,35) | 1604,73 (15 | ,02) 9129,59 | |
E. Азиатский Азиатский | Женский | 84, 74, 84 | 159.94 (6,23) | 32,63 (2,94) | 34,35 (2,02) | 36,60 (1,93) | 48,76 (2,03) | 43,41 (2,24) | 14,47 (1,11) | 125) 451292 (100,99) 4,77) | ||
| N.Европейский | Мужской | 91, 91, 91 | 181,00 (7,72) | 26,71 (2,68) | 34,29 (2,42) | 35,66 (2,53) | ) 51,59 | 47,82 (2,78) | 17,62 (1,25) | 1758. 72 (153,36) 72 (153,36) | 54,46 (5,97) | |
| N.Европейский | Женский | 145, 144, 145 | 167,15 (6,70) | 28,19 (2,60) | 30,91 (2,07) | 1,) | 48,62 (2,11) | 44,78 (2,37) | 16,43 (1,02) | 1470,61 (118,68) | 45,80 (5,13) | |
| S. 6,13 (173,91) | 38,50 (5,82) | 37. 47 (2,59) 47 (2,59) | 37,77 (2,57) | 50,81 (2,34) | 46,59 (2,35) | 16,60 (1,23) | 1719,14 (134,48) | 60,83 (8,87) | ||||
| S. Женский | 31, 26, 31 | 157,74 (6,72) | 39,89 (4,72) | 33,20 (1,89) | 34,33 (1,92) | 47,31 (2,13) | 43,57 (2,47) | 7 | 1422,48 (122,86) | 48,52 (6,48) |
Тестирование ускоренной дивергенции
Различия в фенотипах могут накапливаться в популяциях просто из-за генетического дрейфа. Чтобы вызвать положительный направленный отбор, нужно продемонстрировать, что вариации между популяциями больше, чем ожидаемые при генетическом дрейфе. Мы использовали Qst (см. Недавний обзор Leinonen и др. [28]), количественный генетический аналог Fst [10,29], чтобы проверить, проявляют ли определенные аспекты формы носа большую дифференциацию между популяциями, чем ожидалось при генетическом дрейфе. в одиночестве. Qst измеряет степень генетической дифференциации количественного признака в популяциях и определяется как:
Чтобы вызвать положительный направленный отбор, нужно продемонстрировать, что вариации между популяциями больше, чем ожидаемые при генетическом дрейфе. Мы использовали Qst (см. Недавний обзор Leinonen и др. [28]), количественный генетический аналог Fst [10,29], чтобы проверить, проявляют ли определенные аспекты формы носа большую дифференциацию между популяциями, чем ожидалось при генетическом дрейфе. в одиночестве. Qst измеряет степень генетической дифференциации количественного признака в популяциях и определяется как:
где σgb2 и σgw2 — компоненты фенотипической дисперсии из-за аддитивных генетических эффектов среди и внутри популяций, соответственно.Было показано, что, в принципе, ожидается, что распределение Qst количественного признака, которое развилось только в результате генетического дрейфа, будет равно Fst нейтральных генетических маркеров [11,30,31]. Это ожидание позволяет сравнивать Qst с Fst, чтобы проверить, достаточно ли одного генетического дрейфа для объяснения дивергенции признака среди популяций. Если Qst признака в наборе популяций намного больше, чем Fst, это означает, что фенотипическая дифференциация превышает ожидание при нейтральности.
Если Qst признака в наборе популяций намного больше, чем Fst, это означает, что фенотипическая дифференциация превышает ожидание при нейтральности.
Компоненты аддитивной генетической изменчивости, σgb2 и σgw2, обычно оцениваются на основе экспериментов «общего сада», в которых можно контролировать влияние окружающей среды [28]. Это часто невозможно для немодельных организмов, особенно для людей, из-за практических и этических ограничений. В таких случаях σgb2 и σgw2 могут быть оценены на основе межпопуляционных и внутрипопуляционных компонентов фенотипической дисперсии , σpb2 и σpw2, соответственно, при условии, что наследуемость, лежащая в основе этих компонентов, известна:
Qst = cσpb2cσpb2 + 2h3σpw2 = ch3σpb2ch3σpb2 + 2σpw2
(2)
Здесь σpb2 и σpw2 относятся к внутрипопуляционным и внутрипопуляционным компонентам фенотипической дисперсии, а c 83 и h — пропорции c 60 и h. σpb2 и σpw2, соответственно, обусловленные аддитивными генетическими эффектами. И c , и h 2 могут варьироваться от 0 (ни одна из вариаций не обусловлена аддитивными генетическими эффектами) до 1 (вся вариация обусловлена аддитивными генетическими эффектами). Уравнение (2) показывает, что Qst, вычисленное из компонентов фенотипической дисперсии, зависит от соотношения между c и h 2 [32]. Без предварительной информации разумно предположить, что ц / ч 2 = 1, т.е. доля фенотипической дисперсии, обусловленная аддитивными генетическими эффектами, одинакова среди популяций и внутри популяций.Рассчитанный таким образом Qst иногда называют Pst [33]. Однако мы продолжим использовать термин Qst, чтобы избежать путаницы, и оценим справедливость предположения, что c / h 2 = 1 в следующем разделе.
И c , и h 2 могут варьироваться от 0 (ни одна из вариаций не обусловлена аддитивными генетическими эффектами) до 1 (вся вариация обусловлена аддитивными генетическими эффектами). Уравнение (2) показывает, что Qst, вычисленное из компонентов фенотипической дисперсии, зависит от соотношения между c и h 2 [32]. Без предварительной информации разумно предположить, что ц / ч 2 = 1, т.е. доля фенотипической дисперсии, обусловленная аддитивными генетическими эффектами, одинакова среди популяций и внутри популяций.Рассчитанный таким образом Qst иногда называют Pst [33]. Однако мы продолжим использовать термин Qst, чтобы избежать путаницы, и оценим справедливость предположения, что c / h 2 = 1 в следующем разделе.
Мы рассчитали Qst для каждого аспекта формы носа, описанного в предыдущем разделе, по четырем группам населения: i) западноафриканский (N = 40), ii) североевропейский (N = 236), iii) восточноазиатский (N = 40). = 127), и iv) Южная Азия (N = 73) (см. «Методы для критериев отбора»).Мы использовали непараметрический подход начальной загрузки для создания эмпирических распределений Qst и Fst и для проверки того, превышает ли наблюдаемое значение Qst значение Fst (методы). Мы использовали статистику Qst – Fst, которая, согласно нулевой гипотезе генетического дрейфа, должна быть равна нулю. Чем больше Qst – Fst фенотипа, тем сильнее доказательство того, что вариации фенотипа в популяциях больше, чем ожидаемые при одном только генетическом дрейфе. Мы называем выбросы в нейтральном распределении сигналами ускоренной дивергенции для краткости.Сила доказательства ускоренной дивергенции может быть измерена с помощью эмпирического p-значения, которое представляет собой долю бутстрепных значений Qst – Fst, которые меньше нуля. Чтобы сравнить с другими количественными признаками на полигенной основе, мы также проверили, являются ли рост и пигментация кожи сигналами ускоренной дивергенции. Результаты проиллюстрированы, а p-значения перечислены в.
= 127), и iv) Южная Азия (N = 73) (см. «Методы для критериев отбора»).Мы использовали непараметрический подход начальной загрузки для создания эмпирических распределений Qst и Fst и для проверки того, превышает ли наблюдаемое значение Qst значение Fst (методы). Мы использовали статистику Qst – Fst, которая, согласно нулевой гипотезе генетического дрейфа, должна быть равна нулю. Чем больше Qst – Fst фенотипа, тем сильнее доказательство того, что вариации фенотипа в популяциях больше, чем ожидаемые при одном только генетическом дрейфе. Мы называем выбросы в нейтральном распределении сигналами ускоренной дивергенции для краткости.Сила доказательства ускоренной дивергенции может быть измерена с помощью эмпирического p-значения, которое представляет собой долю бутстрепных значений Qst – Fst, которые меньше нуля. Чтобы сравнить с другими количественными признаками на полигенной основе, мы также проверили, являются ли рост и пигментация кожи сигналами ускоренной дивергенции. Результаты проиллюстрированы, а p-значения перечислены в. Мы относимся к фенотипам, прошедшим строгую поправку Бонферонни (значение p <0,0071 = 0,05 / 7 для семи признаков формы носа), как к признакам ускоренного расхождения между популяциями.
Мы относимся к фенотипам, прошедшим строгую поправку Бонферонни (значение p <0,0071 = 0,05 / 7 для семи признаков формы носа), как к признакам ускоренного расхождения между популяциями.
Бутстрепное распределение Qst – Fst для каждого фенотипа (показанное графиком скрипки) сравнивается с ожидаемым значением нуля при нейтральности (горизонтальная пунктирная линия). Фенотипы, которые демонстрируют ускоренную дивергенцию (с использованием порогового значения p, скорректированного по Бонферронни, равного 0,0071), показаны красным.
Таблица 2
Результаты тестов ускоренной дивергенции между популяциями.
| Фенотип | Qst c | эмпирическое значение p d | Критическое значение c / h 2 e | Высота 912. 177 | 0,1433 | > 1 | |
|---|---|---|---|---|---|---|---|
| Пигментация кожи | 0,642 | 0,0001 | 0,25 | ||||
| Ширина края | 0,467 | 0,0028 | 0,0049 | 0,55 | |||
| Высота носа | 0,023 | 0,7886 | > 1 | ||||
| Длина носового гребня | 0,035 | 0. 7053 7053 | > 1 | ||||
| Выступ кончика носа | 0,306 | 0,0274 | ~ 1 | ||||
| Площадь внешней поверхности | 0,127 | 0,2601 | 0,1919 | > 1 |
Как и ожидалось, пигментация кожи свидетельствует об ускорении дивергенции между популяциями (Qst = 0,642, значение p = 1,00E-04). Это соответствует идее о том, что различия в пигментации кожи, по крайней мере, среди континентальных популяций, были вызваны положительным отбором. Высота, с другой стороны, не показывает сигналов об ускоренной дивергенции (Qst = 0,177, p-value = 0,14). Хотя рост, по-видимому, находится под положительным отбором в пределах европейских и африканских популяций отдельно [19,34], он, по-видимому, не вызван дивергентным давлением отбора в рассматриваемых здесь популяциях. Из всех измерений формы носа только ширина ноздри (Qst = 0,467, значение p = 2,80E-03) и ширина основания носа (Qst = 0,440, значение p = 4,90E-03) показывают сигналы ускоренной дивергенции.Однако степень расхождения этих признаков не так высока, как наблюдаемая для пигментации кожи.
Высота, с другой стороны, не показывает сигналов об ускоренной дивергенции (Qst = 0,177, p-value = 0,14). Хотя рост, по-видимому, находится под положительным отбором в пределах европейских и африканских популяций отдельно [19,34], он, по-видимому, не вызван дивергентным давлением отбора в рассматриваемых здесь популяциях. Из всех измерений формы носа только ширина ноздри (Qst = 0,467, значение p = 2,80E-03) и ширина основания носа (Qst = 0,440, значение p = 4,90E-03) показывают сигналы ускоренной дивергенции.Однако степень расхождения этих признаков не так высока, как наблюдаемая для пигментации кожи.
Оценка достоверности предположений относительно
c и h 2 Мы оценили Qst в предположении, что c / h 2 = 1. Другими словами, мы предполагаем, что соотношение межпопуляционной дисперсии признака из-за аддитивных генетических эффектов такая же, как и доля внутрипопуляционной дисперсии из-за аддитивных генетических эффектов. Это может быть неверным предположением, если большая часть фенотипической дисперсии между популяциями обусловлена прямым воздействием окружающей среды (т.е. фенотипической пластичностью). Если истинное значение ц / ч 2 значительно ниже 1, полученные по фенотипу оценки генетической дисперсии между популяциями и, следовательно, Qst будут завышены, что приведет к ложноположительным выводам относительно роли отбора в управлении фенотипической дифференциацией. Этот факт в значительной степени игнорировался в предыдущих исследованиях, в которых подходы на основе Qst использовались для изучения краниофациальной дивергенции [12–14,35].В этих исследованиях h 2 было принято равным 0,55, а c неявно предполагалось равным 1, в результате чего c / h 2 было больше 1. Хотя это могло быть верно для некоторых признаков (например, см. пигментацию кожи в), это, по нашему мнению, антиконсервативный подход, поскольку он предполагает, что наследуемость между популяциями выше, чем наследуемость внутри популяций.
Это может быть неверным предположением, если большая часть фенотипической дисперсии между популяциями обусловлена прямым воздействием окружающей среды (т.е. фенотипической пластичностью). Если истинное значение ц / ч 2 значительно ниже 1, полученные по фенотипу оценки генетической дисперсии между популяциями и, следовательно, Qst будут завышены, что приведет к ложноположительным выводам относительно роли отбора в управлении фенотипической дифференциацией. Этот факт в значительной степени игнорировался в предыдущих исследованиях, в которых подходы на основе Qst использовались для изучения краниофациальной дивергенции [12–14,35].В этих исследованиях h 2 было принято равным 0,55, а c неявно предполагалось равным 1, в результате чего c / h 2 было больше 1. Хотя это могло быть верно для некоторых признаков (например, см. пигментацию кожи в), это, по нашему мнению, антиконсервативный подход, поскольку он предполагает, что наследуемость между популяциями выше, чем наследуемость внутри популяций. Во всяком случае, противоположный случай ( c / h 2 <1) более правдоподобен, поскольку мы ожидаем, что экологические вариации будут большими среди географически удаленных популяций.Имея это в виду, мы оценили чувствительность результатов Qst – Fst к случаю, когда c / h 2 <1 [32], определив «критическое значение» c / h 2 , при котором 95% нижняя граница Qst соответствует 95% верхней границе Fst (). Меньшие критические значения означают, что сравнения Qst – Fst на основе фенотипа устойчивы к предположениям ц / ч 2 . Такие «анализы чувствительности», хотя и распространены в литературе по молекулярной экологии, недостаточно представлены в генетических и антропологических исследованиях человека.показывает кривые чувствительности к особенностям роста, пигментации кожи и формы носа. Пигментация кожи, которая показывает очень высокое значение Qst, имеет критическое значение 0,25, что намного ниже 1 (и).
Во всяком случае, противоположный случай ( c / h 2 <1) более правдоподобен, поскольку мы ожидаем, что экологические вариации будут большими среди географически удаленных популяций.Имея это в виду, мы оценили чувствительность результатов Qst – Fst к случаю, когда c / h 2 <1 [32], определив «критическое значение» c / h 2 , при котором 95% нижняя граница Qst соответствует 95% верхней границе Fst (). Меньшие критические значения означают, что сравнения Qst – Fst на основе фенотипа устойчивы к предположениям ц / ч 2 . Такие «анализы чувствительности», хотя и распространены в литературе по молекулярной экологии, недостаточно представлены в генетических и антропологических исследованиях человека.показывает кривые чувствительности к особенностям роста, пигментации кожи и формы носа. Пигментация кожи, которая показывает очень высокое значение Qst, имеет критическое значение 0,25, что намного ниже 1 (и).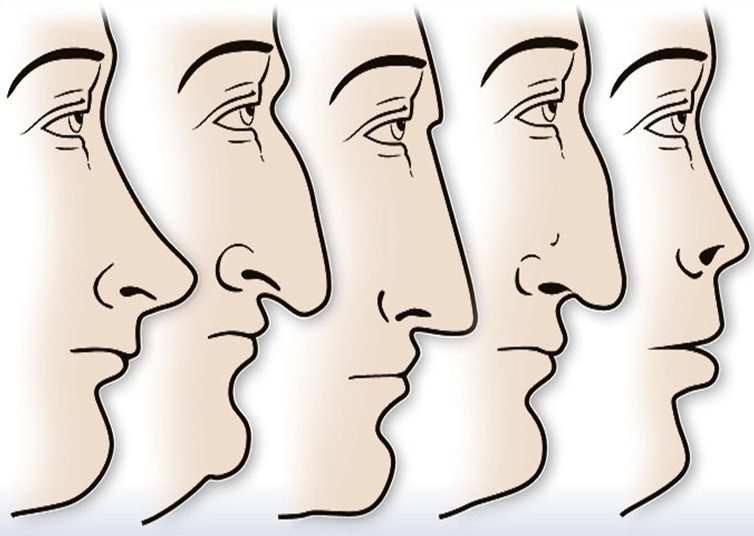 Критическое значение ширины ноздрей составляет 0,50, а ширина основания крыла — 0,55, что также намного меньше 1 (и). Таким образом, мы ожидаем, что эти фенотипы будут демонстрировать сигналы ускоренной дивергенции, даже если истинные значения c / h 2 были намного меньше единицы.
Критическое значение ширины ноздрей составляет 0,50, а ширина основания крыла — 0,55, что также намного меньше 1 (и). Таким образом, мы ожидаем, что эти фенотипы будут демонстрировать сигналы ускоренной дивергенции, даже если истинные значения c / h 2 были намного меньше единицы.
Изменение медианы Qst роста, пигментации кожи и формы носа в зависимости от c / h 2 показано сплошной красной линией. Медиана Fst показана сплошной синей линией. Нижняя и верхняя 95% границы для Qst и Fst показаны пунктирными красными и синими линиями соответственно. Критическое значение, при котором нижняя граница Qst встречается с верхней границей Fst, показано в верхнем левом углу каждого графика.Более низкие критические значения указывают на то, что Qst – Fst более устойчив к предположениям c / h 2 .
Таблица 3
Наследственность фенотипов внутри и между популяциями.
| Фенотип | Размер выборки (для h г 2 ) — Европейцы f | h g 2 (SE) | Размер образца (для h y 2 ) –Смешанный европейский и W.Африканский г | h y 2 (SE) | ||||
|---|---|---|---|---|---|---|---|---|
| Высота | 1825 | 0,394 (0,174) | 409 | Индекс меланина | 1,231 | 0,191 (0,263) | NA | NA |
| Ширина ноздрей | 1,718 | 0,504 (0,187) | 409 | 1,718 | 0. 481 (0,188) 481 (0,188) | 409 | 0,212 (0,093) | |
| Высота носа | 1,718 | 0,441 (0,186) | 409 | 0,030 (0,076) | Длина гребня | (0,188)409 | 0,059 (0,078) | |
| Выступ кончика носа | 1,718 | 0,401 (0,191) | 409 | 0,177 (0,088) | 1 792 | . | 409 | 0,121 (0,086) |
| Область ноздри | 1,718 | 0,657 (0,187) | 409 | 0,059 (0,088) |
 449 (0,187)
449 (0,187)среди популяций
В дополнение к анализу чувствительности мы хотели продемонстрировать, что различия в форме носа у людей как внутри, так и между популяциями являются наследственными. Внутрипопуляционная наследственность ( h 2 ) по фенотипу традиционно оценивается на основе данных, собранных на больших группах близнецов или родословных, в которых известны генетические отношения между особями [36].Недавно Янг и его коллеги (2010) представили линейный подход смешанной модели, который можно использовать для оценки альтернативной статистики у неродственных людей: доля фенотипической дисперсии, объясняемая генотипированными SNP ( h g 2 ) [18]. Основное преимущество этого подхода перед традиционными подходами, основанными на близнецах и родословных, заключается в том, что он применим к данным от неродственных лиц, которые более доступны и легче собирать в больших количествах.Мы вычислили h g 2 как оценку h 2 для роста, индекса меланина и формы носа, используя 118 420 аутосомных SNP в выборке неродственных людей европейского происхождения. , реализованный в программе GCTA [37] (Методы). Значения h g 2 представлены в. Наследственность по росту у европейцев ( h g 2 = 0.394; N = 1,825) аналогична ранее опубликованным оценкам [18]. Интересно, что наследуемость пигментации кожи у европейцев очень низкая ( h g 2 = 0,191; N = 1231), что, по нашему мнению, можно объяснить снижением генетической изменчивости локусов пигментации кожи из-за сильный положительный отбор для более светлой кожи у европейцев [17,21,38]. Почти все аспекты формы носа кажутся европейцам очень наследуемыми ( h g 2 диапазон: 0.401–0,657 N = 1,718) ().
Основное преимущество этого подхода перед традиционными подходами, основанными на близнецах и родословных, заключается в том, что он применим к данным от неродственных лиц, которые более доступны и легче собирать в больших количествах.Мы вычислили h g 2 как оценку h 2 для роста, индекса меланина и формы носа, используя 118 420 аутосомных SNP в выборке неродственных людей европейского происхождения. , реализованный в программе GCTA [37] (Методы). Значения h g 2 представлены в. Наследственность по росту у европейцев ( h g 2 = 0.394; N = 1,825) аналогична ранее опубликованным оценкам [18]. Интересно, что наследуемость пигментации кожи у европейцев очень низкая ( h g 2 = 0,191; N = 1231), что, по нашему мнению, можно объяснить снижением генетической изменчивости локусов пигментации кожи из-за сильный положительный отбор для более светлой кожи у европейцев [17,21,38]. Почти все аспекты формы носа кажутся европейцам очень наследуемыми ( h g 2 диапазон: 0.401–0,657 N = 1,718) ().
Оценить наследуемость среди популяций ( c ) сложнее, поскольку генетические эффекты между географически удаленными популяциями могут быть смешаны с большими экологическими эффектами. Этот параметр часто можно надежно оценить только с помощью экспериментов «общего сада», в которых могут быть приняты во внимание систематические различия в окружающей среде, которые могут быть смешаны с генетическими различиями. Однако, как упоминалось ранее, этот подход возможен только в модельных организмах, которые поддаются экспериментальным манипуляциям.У людей и других немодельных организмов это серьезное ограничение, которое можно преодолеть, изучая встречающиеся в природе смешанные популяции.
Процесс смешения объединяет генофонды двух или более популяций, которые могли разойтись из-за генетического дрейфа, мутации и отбора. Смешивание, которое может происходить неоднократно в течение нескольких поколений, с последующей рекомбинацией, приводит к хромосомам, которые по сути представляют собой мозаику сегментов предков () [39]. В случайно спаривающейся смешанной популяции сегменты родословной разделяются случайным образом по отношению к окружающей среде, что отделяет межпопуляционные генетические эффекты на фенотип от эффектов окружающей среды, что позволяет оценить генетическую вариативность, лежащую в основе фенотипа.Мы предполагаем, что метод, недавно разработанный для оценки наследуемости в смешанных популяциях (Zaitlen et al . (2014)) [40], может дать достоверную оценку c . Зайтлен и др. . (2014) расширяют метод, разработанный Янгом и др. . (2010) [18], используя локальную родословную в SNP вместо генотипов, чтобы построить матрицу генетических отношений между смешанными особями. Доля фенотипической дисперсии в смешанной популяции, которая может быть объяснена местным происхождением ( h y 2 ), концептуально эквивалентна c между родительскими популяциями.
Смешивание, которое может происходить неоднократно в течение нескольких поколений, с последующей рекомбинацией, приводит к хромосомам, которые по сути представляют собой мозаику сегментов предков () [39]. В случайно спаривающейся смешанной популяции сегменты родословной разделяются случайным образом по отношению к окружающей среде, что отделяет межпопуляционные генетические эффекты на фенотип от эффектов окружающей среды, что позволяет оценить генетическую вариативность, лежащую в основе фенотипа.Мы предполагаем, что метод, недавно разработанный для оценки наследуемости в смешанных популяциях (Zaitlen et al . (2014)) [40], может дать достоверную оценку c . Зайтлен и др. . (2014) расширяют метод, разработанный Янгом и др. . (2010) [18], используя локальную родословную в SNP вместо генотипов, чтобы построить матрицу генетических отношений между смешанными особями. Доля фенотипической дисперсии в смешанной популяции, которая может быть объяснена местным происхождением ( h y 2 ), концептуально эквивалентна c между родительскими популяциями. Мы рассуждаем о том, что на генотипическом уровне генетическая изменчивость в смешанной популяции должна быть суммой генетической изменчивости в родительских популяциях и генетической изменчивости между ними. Вариации в масштабе местной родословной представляют только генетические различия между популяциями (). Таким образом, доля фенотипической изменчивости, которая может быть объяснена местным происхождением ( h y 2 ), должна быть эквивалентна c .
Мы рассуждаем о том, что на генотипическом уровне генетическая изменчивость в смешанной популяции должна быть суммой генетической изменчивости в родительских популяциях и генетической изменчивости между ними. Вариации в масштабе местной родословной представляют только генетические различия между популяциями (). Таким образом, доля фенотипической изменчивости, которая может быть объяснена местным происхождением ( h y 2 ), должна быть эквивалентна c .
Генетическая изменчивость между двумя родительскими популяциями представлена различием в цвете хромосом, тогда как генетическая изменчивость внутри популяций представлена интенсивностью оттенка. Смесь объединяет генетические вариации обеих популяций. На генотипическом уровне генетическая изменчивость в смешанных популяциях состоит как из внутрипопуляционных, так и из межпопуляционных вариаций. Различия на уровне предков — это вариации между двумя родительскими популяциями.
Различия на уровне предков — это вариации между двумя родительскими популяциями.
Следуя этим рассуждениям, мы оценили h y 2 от местного происхождения на 623 625 аутосомных SNP в выборке из 409 жителей Кабо-Верде, которые произошли в основном от западноафриканцев и европейцев (см. подробности и результаты). показывает, что фенотипические различия между западными африканцами и европейцами по росту ( h y 2 = 0.224, N = 409) вполне по наследству. Результаты для данных о пигментации кожи здесь не представлены, так как они были недоступны. Различия во многих аспектах формы носа также наследуются (ширина ноздри: h y 2 = 0,226, ширина основания носа: h y 2 = 0,212, Выступ кончика носа: 0,177, Площадь внешней поверхности: h y 2 = 0. 121, N = 409). Другие аспекты формы носа могут не передаваться по наследству между этими двумя популяциями (высота носа: h y 2 = 0,03, h y 2 = 0,059, N = 409). Еще одно интересное наблюдение заключается в том, что оценки h y 2 обычно ниже, чем оценки h g 2 для всех признаков.Это предполагает, что для большинства человеческих черт аддитивная генетическая дисперсия между популяциями может быть меньше аддитивной генетической дисперсии внутри популяций, что согласуется с тем фактом, что большая часть генетической изменчивости у людей существует внутри популяций. Тем не менее, мы осторожно переоцениваем этот вывод, поскольку наследственность была оценена только у людей с западноафриканским и европейским происхождением и не отражала вариации внутри и между другими популяциями.
121, N = 409). Другие аспекты формы носа могут не передаваться по наследству между этими двумя популяциями (высота носа: h y 2 = 0,03, h y 2 = 0,059, N = 409). Еще одно интересное наблюдение заключается в том, что оценки h y 2 обычно ниже, чем оценки h g 2 для всех признаков.Это предполагает, что для большинства человеческих черт аддитивная генетическая дисперсия между популяциями может быть меньше аддитивной генетической дисперсии внутри популяций, что согласуется с тем фактом, что большая часть генетической изменчивости у людей существует внутри популяций. Тем не менее, мы осторожно переоцениваем этот вывод, поскольку наследственность была оценена только у людей с западноафриканским и европейским происхождением и не отражала вариации внутри и между другими популяциями. В целом наши результаты показывают, что генетические различия лежат в основе различий во многих аспектах формы носа как внутри, так и между популяциями.
В целом наши результаты показывают, что генетические различия лежат в основе различий во многих аспектах формы носа как внутри, так и между популяциями.
Тестирование на адаптацию к климату
Ранее в нескольких исследованиях было показано, что формы носового отверстия и носовой полости коррелируют с климатическими переменными, связанными с температурой и влажностью, так что у людей из холодно-сухого климата носовые полости более узкие, чем у людей. из теплого влажного климата [41,42]. Мы были заинтересованы в том, чтобы проверить, демонстрируют ли аспекты формы внешнего носа, демонстрирующие необычно высокую дифференциацию среди популяций, на основе анализа Qst – Fst, корреляции с климатом.Для этой цели мы выбрали из подмножества, использованного в анализе Qst, женщин с генетическими данными, родители которых родились в регионе, совпадающем с их континентальной родословной (N = 140) (). Это было сделано для того, чтобы присвоить каждому человеку климатическую ценность, наиболее близкую к климату их «предков». Поскольку у нас не было данных о генотипах мужчин североевропейского происхождения, мы использовали только женщин для этого анализа. Данные генотипа были необходимы для корректировки генетической структуры (см. Методы).Мы проверили корреляцию ширины ноздрей и ширины крылового основания с тремя климатическими переменными: i) среднегодовой температурой (далее именуемой температурой), ii) относительной влажностью и iii) абсолютной влажностью. Выбор этих климатических переменных следует из функционального значения носа в нагревании и увлажнении вдыхаемого воздуха. Мы также проверили, связана ли пигментация кожи с уровнем УФ-В. Это было использовано в качестве доказательства принципа, поскольку многочисленные свидетельства предполагают, что различия в пигментации кожи в разных популяциях развивались в первую очередь в ответ на ультрафиолетовое излучение [17, 20–22].Связь между фенотипами и климатическими переменными была проверена с использованием линейных смешанных моделей, которые корректируют возраст, ИМТ и генетическое сходство (методы).
Поскольку у нас не было данных о генотипах мужчин североевропейского происхождения, мы использовали только женщин для этого анализа. Данные генотипа были необходимы для корректировки генетической структуры (см. Методы).Мы проверили корреляцию ширины ноздрей и ширины крылового основания с тремя климатическими переменными: i) среднегодовой температурой (далее именуемой температурой), ii) относительной влажностью и iii) абсолютной влажностью. Выбор этих климатических переменных следует из функционального значения носа в нагревании и увлажнении вдыхаемого воздуха. Мы также проверили, связана ли пигментация кожи с уровнем УФ-В. Это было использовано в качестве доказательства принципа, поскольку многочисленные свидетельства предполагают, что различия в пигментации кожи в разных популяциях развивались в первую очередь в ответ на ультрафиолетовое излучение [17, 20–22].Связь между фенотипами и климатическими переменными была проверена с использованием линейных смешанных моделей, которые корректируют возраст, ИМТ и генетическое сходство (методы). Пол не был включен в качестве ковариаты, поскольку использовались только самки. Мы использовали тест отношения правдоподобия (LRT), чтобы оценить статистическую значимость наклона между фенотипом и климатической переменной. Статистика LRT и соответствующие ей p-значения были получены путем сравнения полной (включая предсказатель климата) и сокращенной (предсказатель климата удален) моделей.Результаты представлены в.
Пол не был включен в качестве ковариаты, поскольку использовались только самки. Мы использовали тест отношения правдоподобия (LRT), чтобы оценить статистическую значимость наклона между фенотипом и климатической переменной. Статистика LRT и соответствующие ей p-значения были получены путем сравнения полной (включая предсказатель климата) и сокращенной (предсказатель климата удален) моделей.Результаты представлены в.
Отдельные точки обозначают места рождения родителей с линией, соединяющей двух родителей. Единственная точка означает, что оба родителя родились в одном месте. Значения климата в этих местах использовались для проверки сигналов об адаптации к климату.
Таблица 4
Результаты теста на адаптацию к климату.
| Пигментация кожи m (N = 126) | |||||||||||||
| Наклон h | SE 912 T j | LRT k | P-значение l | ||||||||||
| UVB | 3. 16E-03 16E-03 | 5.09E-04 | 6.23 | 55.20 | 1.09E-13 | ||||||||
| Ширина ноздрей (N = 140) | |||||||||||||
| Наклон | T | LRT | Значение P | ||||||||||
| Темп. -3.626 | 2,192 | -1,65 | 2,71 | 0,100 | |||||||||
| Абсолютная влажность | 0,149 | 0,054 | 2,73 | -7,28 | = 140) | ||||||||
| Наклон | SE | T | LRT | P-value | |||||||||
Темп. 065 065 | 0,033 | 1,97 | 3,82 | 0,051 | |||||||||
| Относительная влажность | -1,642 | 2,083 | -0,79 | 0,62 | -0,79 | 0,62 92 | 0,49 | 0,62 | 0,49 | 0,052 | 2,09 | 4,30 | 0,038 |
Как и ожидалось, мы видим сильную взаимосвязь между пигментацией кожи и УФ-В (N = 126, T = 6. 23, LRT = 55.20, p-значение = 1.09E-13). Положительные значения наклона и t-статистики указывают на то, что люди, как правило, темнее в регионах с более высоким воздействием УФ-В и наоборот. Эта закономерность дополнительно подтверждает идею о том, что адаптация к ультрафиолетовому излучению сыграла важную роль в эволюции пигментации кожи у людей. Ширина ноздрей значительно коррелирует с температурой (N = 140, T = 3,09, LRT = 9,24, p-value = 2,37E-03) и абсолютной влажностью (N = 140, T = 2,73, LRT = 7.28, p-значение = 6.97E-03). Положительные наклоны указывают на то, что люди из теплого влажного климата в среднем обычно имеют более широкую ноздрю, тогда как люди из прохладно-сухого климата, как правило, имеют более узкую ноздрю. Это согласуется с ранее описанными паттернами экогеографических вариаций ширины носовой апертуры и носовой полости [41,42]. Ширина ноздрей не имела значимой корреляции с относительной влажностью (N = 140, T = -1,65, LRT = 2,71, значение p = 0,100). Это неудивительно, поскольку абсолютная влажность более важна для физиологического функционирования носа, чем относительная влажность [43].
23, LRT = 55.20, p-значение = 1.09E-13). Положительные значения наклона и t-статистики указывают на то, что люди, как правило, темнее в регионах с более высоким воздействием УФ-В и наоборот. Эта закономерность дополнительно подтверждает идею о том, что адаптация к ультрафиолетовому излучению сыграла важную роль в эволюции пигментации кожи у людей. Ширина ноздрей значительно коррелирует с температурой (N = 140, T = 3,09, LRT = 9,24, p-value = 2,37E-03) и абсолютной влажностью (N = 140, T = 2,73, LRT = 7.28, p-значение = 6.97E-03). Положительные наклоны указывают на то, что люди из теплого влажного климата в среднем обычно имеют более широкую ноздрю, тогда как люди из прохладно-сухого климата, как правило, имеют более узкую ноздрю. Это согласуется с ранее описанными паттернами экогеографических вариаций ширины носовой апертуры и носовой полости [41,42]. Ширина ноздрей не имела значимой корреляции с относительной влажностью (N = 140, T = -1,65, LRT = 2,71, значение p = 0,100). Это неудивительно, поскольку абсолютная влажность более важна для физиологического функционирования носа, чем относительная влажность [43].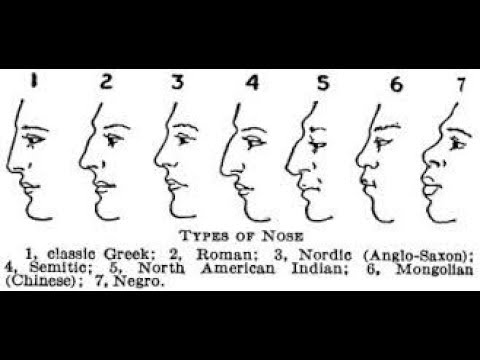 Ширина основания крыльев существенно не коррелирует ни с температурой (N = 140, T = 1,97, LRT = 3,82, значение p = 0,051), ни с относительной влажностью (N = 140, T = -0,79, LRT = 0,62, значение p = 0,431) и лишь слегка коррелирует с абсолютной влажностью (N = 140, T = 2,09, LRT = 4,30, p-значение = 0,038). Это может означать, что не ширина носа как таковая , а ширина ноздрей находится под давлением отбора. Однако даже ширина ноздрей не так сильно коррелирует с температурой и влажностью, как пигментация кожи — с УФ-В, что позволяет предположить, что сила отбора, оказываемого температурой на ширину носа, вероятно, будет намного слабее, чем давление отбора, оказываемое воздействием УФ-В на пигментацию кожи. .
Ширина основания крыльев существенно не коррелирует ни с температурой (N = 140, T = 1,97, LRT = 3,82, значение p = 0,051), ни с относительной влажностью (N = 140, T = -0,79, LRT = 0,62, значение p = 0,431) и лишь слегка коррелирует с абсолютной влажностью (N = 140, T = 2,09, LRT = 4,30, p-значение = 0,038). Это может означать, что не ширина носа как таковая , а ширина ноздрей находится под давлением отбора. Однако даже ширина ноздрей не так сильно коррелирует с температурой и влажностью, как пигментация кожи — с УФ-В, что позволяет предположить, что сила отбора, оказываемого температурой на ширину носа, вероятно, будет намного слабее, чем давление отбора, оказываемое воздействием УФ-В на пигментацию кожи. .
Следуя предложению одного из обозревателей, мы исследовали, были ли наблюдаемые корреляции фенотип-климат обусловлены какой-либо одной популяцией. Для этого мы повторно оценили наклоны фенотипа-климата (пигментация кожи в зависимости от УФ-В и ширины ноздрей в зависимости от температуры) после удаления каждой популяции по очереди. показывает, что 95% доверительные интервалы наклона перекрываются с нулем (красная линия внутри) только тогда, когда северные европейцы удалены, предполагая, что обе корреляции фенотип-климат обусловлены в первую очередь включением N.Европейцы. Этот эффект может указывать на положительный отбор для более светлой пигментации кожи и более узких носовых ходов в популяциях более высоких широт. Однако, поскольку наша выборка популяций на более высоких широтах ограничена (), этот результат требует дальнейшего изучения.
показывает, что 95% доверительные интервалы наклона перекрываются с нулем (красная линия внутри) только тогда, когда северные европейцы удалены, предполагая, что обе корреляции фенотип-климат обусловлены в первую очередь включением N.Европейцы. Этот эффект может указывать на положительный отбор для более светлой пигментации кожи и более узких носовых ходов в популяциях более высоких широт. Однако, поскольку наша выборка популяций на более высоких широтах ограничена (), этот результат требует дальнейшего изучения.
Точечная оценка и 95% доверительные интервалы уклонов фенотип-климат после удаления каждой популяции (по оси Y) показаны для A) пигментации кожи и УФ-излучения B, B) ширины и температуры ноздрей и C) ширины ноздрей. и абсолютная влажность.Во всех случаях 95% доверительный интервал наклона перекрывается с нулем (красная вертикальная линия), только когда европейцы удалены.
Обсуждение
Разнообразие черт лица в разных популяциях людей давно интересовало ученых. Несмотря на то, что генетический дрейф играл преобладающую роль в эволюции человека, внешние физические черты, такие как форма лица и пигментация кожи, из-за их близости к окружающей среде, также, вероятно, находились под влиянием естественного отбора.Накоплены существенные доказательства, подтверждающие гипотезу о том, что различия в пигментации кожи в разных популяциях человека эволюционировали в основном в ответ на давление отбора, наложенное воздействием ультрафиолетового излучения [20]. Неясно, как отбор мог повлиять на форму лица — черту, которая также весьма варьируется в разных популяциях, вероятно, потому, что до сих пор ей уделялось гораздо меньше внимания. Учитывая сложность лица, мы выбрали для изучения одну особенно интересную и изменчивую часть лица; нос.Общий вопрос, лежащий в основе этого исследования, таков: сыграла ли адаптация к климату важную роль в изменении формы человеческого носа? Чтобы ответить на этот вопрос, мы сформулировали две гипотезы: i. дивергентный отбор был вовлечен в дифференциацию определенных аспектов формы носа в разных популяциях, и ii.
Несмотря на то, что генетический дрейф играл преобладающую роль в эволюции человека, внешние физические черты, такие как форма лица и пигментация кожи, из-за их близости к окружающей среде, также, вероятно, находились под влиянием естественного отбора.Накоплены существенные доказательства, подтверждающие гипотезу о том, что различия в пигментации кожи в разных популяциях человека эволюционировали в основном в ответ на давление отбора, наложенное воздействием ультрафиолетового излучения [20]. Неясно, как отбор мог повлиять на форму лица — черту, которая также весьма варьируется в разных популяциях, вероятно, потому, что до сих пор ей уделялось гораздо меньше внимания. Учитывая сложность лица, мы выбрали для изучения одну особенно интересную и изменчивую часть лица; нос.Общий вопрос, лежащий в основе этого исследования, таков: сыграла ли адаптация к климату важную роль в изменении формы человеческого носа? Чтобы ответить на этот вопрос, мы сформулировали две гипотезы: i. дивергентный отбор был вовлечен в дифференциацию определенных аспектов формы носа в разных популяциях, и ii. климат является агентом отбора в тех случаях, когда может быть задействован дивергентный отбор.
климат является агентом отбора в тех случаях, когда может быть задействован дивергентный отбор.
Чтобы проверить первую гипотезу, мы использовали сравнения Qst – Fst, чтобы выяснить, больше ли средняя разница в форме носа среди популяций, чем ожидаемая только при генетическом дрейфе.Оценка Qst основана на учете внутрипопуляционных и межпопуляционных компонентов фенотипической дисперсии, обусловленных аддитивными генетическими эффектами. Эти компоненты дисперсии идеально рассчитываются с помощью экспериментов, в которых можно контролировать влияние переменных окружающей среды [28]. Это представляет собой практическую и этическую проблему для немодельных организмов, таких как люди, которые не поддаются требуемому типу экспериментов. Чтобы обойти эти ограничения, исследования часто рассчитывают Qst непосредственно из данных фенотипа в предположении, что наследуемость внутри популяции ( h 2 ) равна наследуемости между популяциями ( c ).Рассчитанный таким образом Qst, вероятно, лучше всего считать «минимальным» Qst, предложенным Релетфордом (1994) [35]. Однако представление о том, что здесь принимается минимальное значение Qst, основывается на том, что h 2 всегда меньше c , что, как отмечалось ранее, является антиконсервативным. Фактически, генетическая изменчивость между популяциями, вероятно, будет меньше, чем генетическая изменчивость внутри популяций для большинства фенотипов. Наш подход аналогичен предыдущим исследованиям в том, что мы также оценили Qst, приняв c = h 2 .Однако мы использовали кривые чувствительности для оценки поведения Qst в случаях, когда c < h 2 . Кроме того, мы демонстрируем, что как внутрипопуляционные, так и межпопуляционные вариации формы носа являются наследственными.
Однако представление о том, что здесь принимается минимальное значение Qst, основывается на том, что h 2 всегда меньше c , что, как отмечалось ранее, является антиконсервативным. Фактически, генетическая изменчивость между популяциями, вероятно, будет меньше, чем генетическая изменчивость внутри популяций для большинства фенотипов. Наш подход аналогичен предыдущим исследованиям в том, что мы также оценили Qst, приняв c = h 2 .Однако мы использовали кривые чувствительности для оценки поведения Qst в случаях, когда c < h 2 . Кроме того, мы демонстрируем, что как внутрипопуляционные, так и межпопуляционные вариации формы носа являются наследственными.
Мы провели сравнения Qst – Fst для семи признаков формы носа: ширины ноздрей, ширины крыловидного основания, высоты носа, длины носового гребня, выступа кончика носа, площади внешней поверхности и площади ноздри.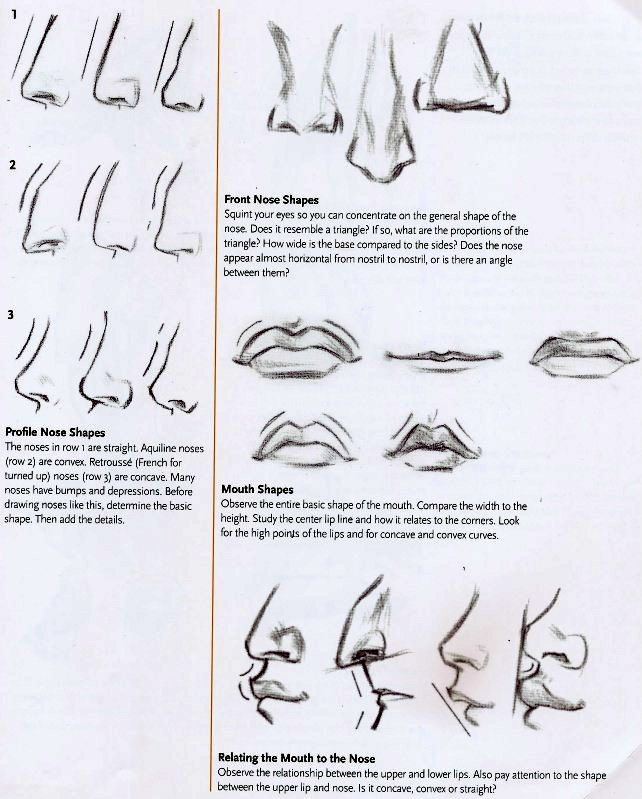 Мы обнаружили, что расхождение в ширине ноздрей и крыловидного основания отклоняется от нейтральных ожиданий, и что эти результаты устойчивы к предположению c = h 2 .Это говорит о том, что ширина ноздрей и крыльев крыльев, возможно, изменилась в разных популяциях из-за дивергентного отбора.
Мы обнаружили, что расхождение в ширине ноздрей и крыловидного основания отклоняется от нейтральных ожиданий, и что эти результаты устойчивы к предположению c = h 2 .Это говорит о том, что ширина ноздрей и крыльев крыльев, возможно, изменилась в разных популяциях из-за дивергентного отбора.
Затем мы выдвинули гипотезу о том, что расхождение этих двух характеристик обусловлено адаптацией к климату. Чтобы исследовать это, мы проверили, коррелирует ли пространственное распределение этих признаков со средней годовой температурой и влажностью. Наши результаты показывают, что ширина ноздрей сильно коррелирует с температурой и абсолютной влажностью. Положительное направление эффектов указывает на то, что более широкие носы чаще встречаются в теплом влажном климате, а более узкие носы чаще встречаются в холодно-сухом климате.Однако ширина ноздрей не связана с относительной влажностью. Это неудивительно, поскольку абсолютная влажность, вероятно, более важна для дыхания, чем относительная влажность [43]. Ширина основания крыла только слабо, если вообще коррелирует с температурой и абсолютной влажностью, предполагая, что сигнал адаптации к климату может быть специфическим для ширины ноздрей. Исследования вычислительной гидродинамики (CFD) показывают, что геометрия носа важна для его дыхательных функций [5,44].Вдыхаемый воздух достигает 90% от требуемых уровней температуры и влажности еще до того, как достигает носоглотки, что делает полость носа, особенно носовые раковины, основным кондиционирующим аппаратом в дыхательных путях [4,5]. Мы также знаем, что геометрия носовых дыхательных путей влияет на скорость вдыхаемого воздуха [4,7,45]. Узкие дыхательные пути в холодно-сухом климате могут способствовать лучшему кондиционированию за счет увеличения турбулентности вдыхаемого воздуха, когда он достигает носовых раковин, тем самым облегчая контакт со слизистой оболочкой носа [5].Тем не менее, мы отмечаем, что область ноздрей не показывает необычно высокой дифференциации между популяциями, что предполагает, что не размер ноздрей, а форма может иметь функциональное значение.
Ширина основания крыла только слабо, если вообще коррелирует с температурой и абсолютной влажностью, предполагая, что сигнал адаптации к климату может быть специфическим для ширины ноздрей. Исследования вычислительной гидродинамики (CFD) показывают, что геометрия носа важна для его дыхательных функций [5,44].Вдыхаемый воздух достигает 90% от требуемых уровней температуры и влажности еще до того, как достигает носоглотки, что делает полость носа, особенно носовые раковины, основным кондиционирующим аппаратом в дыхательных путях [4,5]. Мы также знаем, что геометрия носовых дыхательных путей влияет на скорость вдыхаемого воздуха [4,7,45]. Узкие дыхательные пути в холодно-сухом климате могут способствовать лучшему кондиционированию за счет увеличения турбулентности вдыхаемого воздуха, когда он достигает носовых раковин, тем самым облегчая контакт со слизистой оболочкой носа [5].Тем не менее, мы отмечаем, что область ноздрей не показывает необычно высокой дифференциации между популяциями, что предполагает, что не размер ноздрей, а форма может иметь функциональное значение.
Другая гипотеза, касающаяся функции носа, которой уделялось меньше внимания по сравнению с гипотезой кондиционирования воздуха, — это гипотеза избирательного охлаждения мозга (SBC) [46]. Известно, что некоторые крупные млекопитающие регулируют температуру мозга в гипертермических условиях, часто вызванных длительными упражнениями [47, 48].Эта способность чаще всего встречается у млекопитающих с сетчатой сонной артерией, которая представляет собой сеть капилляров в полости под мозгом, называемой кавернозным синусом. Артериальная кровь, покидающая сетку, в конечном итоге проникает в мозг через Уиллисово кольцо. Венозная кровь, идущая обратно от слизистой оболочки носа, взаимодействует с сеткой сонной артерии в кавернозном синусе и, как полагают, охлаждает кровь, поступающую в мозг у млекопитающих засушливой зоны, таких как Oryx [49], а также у акклиматизированных к зиме животных, таких как как северный олень [50].Хотя люди не обладают такой сеткой и вместо этого имеют единственную артерию, проходящую через кавернозный синус, что заставило некоторых усомниться в роли SBC у людей [51], сторонники утверждают, что сетчатка сонной артерии не является обязательным условием, поскольку SBC присутствует у некоторых животных, у которых нет сетчатки, например у лошадей [52]. Поскольку SBC у людей очень обсуждается, роль различий в форме носа в развитии различий в этом физиологическом процессе, хотя здесь стоит упомянуть, является весьма спекулятивной.
Поскольку SBC у людей очень обсуждается, роль различий в форме носа в развитии различий в этом физиологическом процессе, хотя здесь стоит упомянуть, является весьма спекулятивной.
Климат, возможно, был не единственным фактором, способствовавшим различиям в форме носа у разных популяций. Фактически, мы показываем, что температура слабо коррелирует с шириной ноздрей, особенно по сравнению с корреляцией между пигментацией кожи и УФ-В. Что же тогда может быть селективным агентом, влияющим на изменение формы носа? Ранее мы упоминали, что все аспекты формы носа, изучаемые здесь, являются сексуально диморфными, что вызывает ряд вопросов. Почему существует этот половой диморфизм? Является ли это побочным продуктом циркулирующих гормонов, приводящим к различиям в росте и развитии в раннем взрослом возрасте, или он выполняет адаптивную функцию, например, сигнализирует о сексе другим мужчинам и женщинам? Половой отбор, вероятно, сыграл важную роль в эволюции человека, о чем свидетельствует наличие полового диморфизма во многих физических чертах (например,g. , рост, соотношение талии и бедер, волосы на лице и грудь, и это лишь некоторые из них). Могли ли культурные различия в восприятии доминирования и привлекательности сыграть роль в расхождении формы носа [53]? Могли ли эти представления возникнуть для выбора партнеров, которые сигнализируют об адаптации к местной среде? В самом деле, экологический отбор и половой отбор могут усиливать друг друга, ускоряя фенотипическое расхождение между популяциями, несмотря на продолжающийся поток генов [54]. Это интересные направления исследований, которые необходимо рассмотреть, чтобы составить более полную картину эволюции человеческого носа.
Исследование эволюции формы носа в связи с адаптацией к климату, хотя и интересно с антропологической точки зрения, также имеет медицинское значение. Поскольку люди становятся все более глобальным сообществом, изучение местной адаптации становится все более важным для понимания рисков для здоровья, связанных с жизнью в «чужом» климате. Очевидными примерами таких рисков для здоровья являются повышенный риск солнечных ожогов, рака кожи и дефицита фолиевой кислоты у людей со светлой кожей, подвергающихся высокому воздействию УФ-В, а также низкий вес при рождении и хроническая горная болезнь, связанная с гипоксией на больших высотах [20,55]. Влияет ли морфология внешнего носа или внутренней носовой полости на риск респираторных заболеваний в различных климатических условиях? Пока сложно сказать. Хотя наши результаты подтверждают идею о том, что различия в аспектах формы носа могли развиваться в разных популяциях в результате давления отбора, связанного с климатом, это ранее было продемонстрировано с использованием краниометрических данных [2,12,13,41,42] Отметим, что сигнал об адаптации к климату не очень сильный, особенно по сравнению с пигментацией кожи.Это могло быть связано с более слабым давлением отбора или отбором по изменчивости насаждений, а также из-за представленной здесь разреженной выборки популяций, что является ограничением данного исследования. Эти результаты необходимо будет воспроизвести на более широком наборе групп населения. Мы ожидаем, что исследования, включающие различные группы населения, долгое время жившие в различных средах, таких как тропики, пустыни и приполярные регионы, хорошо восполнят пробелы. Особенно полезным было бы представление населения из высокогорных регионов, таких как жители Анд, тибетцы и эфиопы, которым приходится не только справляться со стрессом из-за холодного и сухого климата, но и из-за низкого уровня кислорода в атмосфере [56].Также было бы информативно изучить нечеловеческих приматов в этом контексте, которые населяют самые разные страны и демонстрируют обширные различия в морфологии носа. Наконец, будущие исследования также должны быть сосредоточены на исследованиях общегеномных ассоциаций (GWAS) для выявления вариантов, влияющих на форму носа. С ростом интереса к идентификации локусов, связанных с формой лица, некоторые GWAS недавно идентифицировали ряд локусов формы носа [57-60]. Генетическая изменчивость в этих локусах будет информативной о природе отбора, а также для определения времени событий отбора.
Влияет ли морфология внешнего носа или внутренней носовой полости на риск респираторных заболеваний в различных климатических условиях? Пока сложно сказать. Хотя наши результаты подтверждают идею о том, что различия в аспектах формы носа могли развиваться в разных популяциях в результате давления отбора, связанного с климатом, это ранее было продемонстрировано с использованием краниометрических данных [2,12,13,41,42] Отметим, что сигнал об адаптации к климату не очень сильный, особенно по сравнению с пигментацией кожи.Это могло быть связано с более слабым давлением отбора или отбором по изменчивости насаждений, а также из-за представленной здесь разреженной выборки популяций, что является ограничением данного исследования. Эти результаты необходимо будет воспроизвести на более широком наборе групп населения. Мы ожидаем, что исследования, включающие различные группы населения, долгое время жившие в различных средах, таких как тропики, пустыни и приполярные регионы, хорошо восполнят пробелы. Особенно полезным было бы представление населения из высокогорных регионов, таких как жители Анд, тибетцы и эфиопы, которым приходится не только справляться со стрессом из-за холодного и сухого климата, но и из-за низкого уровня кислорода в атмосфере [56].Также было бы информативно изучить нечеловеческих приматов в этом контексте, которые населяют самые разные страны и демонстрируют обширные различия в морфологии носа. Наконец, будущие исследования также должны быть сосредоточены на исследованиях общегеномных ассоциаций (GWAS) для выявления вариантов, влияющих на форму носа. С ростом интереса к идентификации локусов, связанных с формой лица, некоторые GWAS недавно идентифицировали ряд локусов формы носа [57-60]. Генетическая изменчивость в этих локусах будет информативной о природе отбора, а также для определения времени событий отбора.
Форма носа была сформирована климатом — ScienceDaily
Большой, маленький, широкий, узкий, длинный или короткий, вздернутый, мопс, крючковатый, выпуклый или выпуклый, люди наследуют форму носа от своих родителей, но в конечном итоге форму По мнению международной группы исследователей, чей-то нос и нос их родителей сформировался в результате длительного процесса адаптации к нашему местному климату.
«Нас интересует недавняя эволюция человека и то, что объясняет очевидные различия в таких вещах, как цвет кожи, цвет волос и само лицо», — сказал Марк Д.Шрайвер, профессор антропологии Пенсильванского университета. «Мы сосредоточились на чертах носа, которые различаются в разных популяциях, и рассмотрели географические различия в отношении температуры и влажности». Сегодня (17 марта) в журнале « PLOS Genetics » исследователи отметили, что «важной функцией носа и носовой полости является кондиционирование вдыхаемого воздуха до того, как он достигнет нижних дыхательных путей».
Они рассматривали различные размеры носа, глядя на ширину ноздрей, расстояние между ноздрями, высоту носа, длину носового гребня, выступ носа, внешнюю область носа и область ноздрей.Измерения проводились с использованием трехмерной визуализации лица.
Различия в человеческом носу могли накапливаться среди популяций с течением времени в результате случайного процесса, называемого генетическим дрейфом. Однако дивергентный отбор — вариация естественного отбора в разных популяциях — также может быть причиной того, что разные популяции имеют разные носы. Разлучить двоих сложно, особенно у людей.
Однако дивергентный отбор — вариация естественного отбора в разных популяциях — также может быть причиной того, что разные популяции имеют разные носы. Разлучить двоих сложно, особенно у людей.
Исследователи обнаружили, что ширина ноздрей и основание носа различались в разных популяциях больше, чем это могло быть объяснено генетическим дрейфом, что указывает на роль естественного отбора в эволюции формы носа у людей.Чтобы показать, что местный климат способствовал этой разнице, исследователи изучили пространственное распределение этих черт и коррелировали их с местными температурами и влажностью. Они показали, что ширина ноздрей сильно коррелирует с температурой и абсолютной влажностью. Исследователи отметили, что «положительное направление эффектов указывает на то, что более широкие носы чаще встречаются в теплом и влажном климате, а более узкие носы чаще встречаются в холодно-сухом климате. климат «.
«Все восходит к Правилу Томпсона (Артур Томпсон), — сказал Шрайвер. «В конце 1800-х годов он сказал, что длинные и тонкие носы встречаются в сухих и холодных областях, а короткие и широкие носы — в жарких и влажных областях. Многие люди проверяли этот вопрос с помощью измерений черепа, но никто не проводил измерения на живые люди «.
«В конце 1800-х годов он сказал, что длинные и тонкие носы встречаются в сухих и холодных областях, а короткие и широкие носы — в жарких и влажных областях. Многие люди проверяли этот вопрос с помощью измерений черепа, но никто не проводил измерения на живые люди «.
Одна из целей носа — кондиционировать вдыхаемый воздух, чтобы он был теплым и влажным. Кажется, что более узкие ноздри изменяют поток воздуха, так что покрытая слизистой оболочкой носа может более эффективно увлажнять и согревать воздух. «Вероятно, это было более важно в холодном и сухом климате», — сказал Шрайвер.Люди с более узкими ноздрями, вероятно, жили лучше и имели больше потомства, чем люди с более широкими ноздрями, в более холодном климате. Это привело к постепенному уменьшению ширины носа у популяций, живущих далеко от экватора.
Шрайвер отмечает, что это не единственное объяснение вариаций формы носа у людей. Исследователи также обнаружили различия между мужчинами и женщинами в чертах носа по всем направлениям. Этот половой диморфизм не является чем-то необычным, поскольку человеческие мужчины, как правило, крупнее человеческих женщин, и их носы тоже будут больше.
Этот половой диморфизм не является чем-то необычным, поскольку человеческие мужчины, как правило, крупнее человеческих женщин, и их носы тоже будут больше.
Он считает, что межпопуляционные различия в размере носа могут возникать по-другому — через половой отбор. Люди могут выбирать себе пару просто потому, что им более симпатичен нос меньшего или большего размера. Если вся группа думает, что чем меньше, тем лучше, то у людей с большим носом будет меньше успеха в воспроизводстве, и в группе будет меньше людей с большим носом. Со временем размер носа в группе уменьшится по сравнению с другими группами, которым отдается предпочтение большим носам. Эти представления о красоте могут быть связаны с тем, насколько хорошо нос приспособлен к местному климату.
По мнению исследователей, экологический отбор и половой отбор могут усиливать друг друга. Однако вопрос о том, была ли эта связь между двумя типами отбора важна для эволюции носа, требует дальнейшего исследования.
История Источник:
Материалы предоставлены Penn State . Оригинал написан А’ндреа Элис Мессер. Примечание. Содержимое можно редактировать по стилю и длине.
Оригинал написан А’ндреа Элис Мессер. Примечание. Содержимое можно редактировать по стилю и длине.
Какие типы носа могут помочь при ринопластике?
Когда дело доходит до эстетических недостатков носа, чаще всего упоминаются его размер, форма, прямолинейность и общая элегантность.В дополнение к этим есть много других причин, по которым люди выбирают ринопластику. Косметическая хирургия носа — одна из самых сложных хирургических процедур на лице. Во-первых, нос — центральная черта лица, и недостатки более заметны, и их труднее скрыть, чем другие недостатки. Не менее важно, что сама ринопластика сложна, потому что требует понимания того, как внешний вид изменится с учетом изменений внутренней костной и хрящевой структуры.
Эстетические недостатки, которые может исправить ринопластика
Ниже приводится список общих эстетических проблем, которые можно улучшить с помощью носовой коррекции:
1. Широкий или плоский носовой мостик
Широкий нос можно сузить, сузив кость в верхней части и / или хрящ в средней и нижней частях. Иногда переносица бывает широкой и плоской и требует увеличения хряща или искусственного материала, чтобы невеста выглядела более элегантной.У разных этнических групп разные характерные формы носа. Увеличение переносицы носа часто встречается у некоторых пациентов из Восточной Азии.
Иногда переносица бывает широкой и плоской и требует увеличения хряща или искусственного материала, чтобы невеста выглядела более элегантной.У разных этнических групп разные характерные формы носа. Увеличение переносицы носа часто встречается у некоторых пациентов из Восточной Азии.
2. Большой выпуклый носовой наконечник
Ринопластика может помочь исправить большой выпуклый кончик носа за счет изменения формы хрящей кончика носа и создания более элегантной треугольной формы. Степень возможного сужения кончика зависит от толщины кожи. У некоторых этнических групп, как правило, более толстая кожа, что ограничивает возможную степень определения кончиков пальцев.
3. Большой или длинный нос
Нос может казаться большим или длинным, или и тем, и другим одновременно. Большой или длинный нос может быть эстетически неприемлемым, так как нарушает гармонию строения лица. Коррекция длинного или большого носа требует сложных техник ринопластики для уменьшения длины (расстояние от места начала носа ниже лба до кончика носа) или выступа (расстояние, на которое нос выступает из лица).
4. Кривой нос
Искривленный нос может быть результатом отклонения перегородки — перегородки, разделяющей правый и левый носовые ходы.Из всех недостатков носа именно этот часто невозможно полностью исправить.
5. Выступающая носовая бугорка или бугорка на переносице
Носовая горбинка может быть наиболее частой причиной, по которой пациенту требуется хирургическое улучшение внешнего вида носа, и из всех недостатков, которые беспокоят пациентов, это тот, который, вероятно, легче всего исправить.
6. «Ненавижу свой профиль, когда улыбаюсь»
Это частая жалоба, обычно связанная с сочетанием перечисленных выше факторов, включая сочетание горбинки, длинного носа и вогнутого кончика носа.Это сочетание анатомических недостатков является одним из самых приятных для хирурга и пациента, потому что послеоперационное улучшение может быть очень значительным.
7. «Мне сделали ринопластику, и я не доволен результатом».