Фенотип внешности: Фенотип человека: Фенотипы европеоидной расы Европы
Содержание
Создана поразительная 3D-реконструкция лиц древних египтян
Художник-криминалист создал трёхмерную реконструкцию внешнего вида трёх мужчин, живших более 2 000 лет назад в Древнем Египте. В качестве основы была использована расшифрованная ДНК трёх древнеегипетских мумий, выделенная из мумифицированных останков.
На цифровой реконструкции мужчины изображены в возрасте 25 лет.
ДНК мумий, найденных в некрополе Абусир эль-Мелек, была секвенирована ещё в 2017 году. Это была первая успешная реконструкция генома древнеегипетской мумии. И не одной, а целых трёх.
Метод, которым воспользовались исследователи, чтобы воссоздать внешность древних египтян, называется ДНК-фенотипирование. Уже из полученных учёными данных художник «собрал» портреты мужчин.
Поясним, что фенотипом называется совокупность внешних и внутренних признаков организма, которые зависят как от генов, так и от случайных мутаций, а также от факторов среды.
Также давно известно, что гены определяют черты лица, цвет волос, кожи и глаз. Более того, учёные давно установили, какие именно гены кодируют те или иные особенности внешности.
Тепловая карта лиц трёх мумий помогла учёным определить различия в их внешности и воссоздать её в мельчайших деталях.
Мумии мужчин были похоронены между 1380 годом до н.э. и 425 годом н.э. Это был первый раз, когда комплексное ДНК-фенотипирование было проведено на основе ДНК столь почтенного возраста.
Учёные установили, что у древних египтян была смуглая кожа, тёмные глаза и волосы. Однако генетически они оказались больше похожи на современных жителей Средиземноморья и Ближнего Востока, чем на современных египтян.
Работать с древней ДНК может быть сложно по двум причинам. Молекулы спустя столько веков могут быть сильно повреждены. Кроме того, обычно в образцах присутствует примесь бактериального генетического материала.
Но учёным и не нужно расшифровывать полный геном каждого отдельного человека, ведь большая часть ДНК идентична у всех людей. Исследователям необходимо проанализировать лишь небольшие участки генома под названием однонуклеотидные полиморфизмы (ОНП). Во многих их них и зашифрованы особенности внешности разных людей.
Когда древняя ДНК не содержит достаточно ОНП, позволяющих составить достаточно точный портрет человека, учёные могут просчитать, какого именно нуклеотида не хватает в имеющейся в наличии цепочке ДНК. Такой статистический прогноз позволяет «заполнить бреши» в геноме и составить наиболее полную картину, в данном случае, внешности человека.
Добавим, что использованный в этой работе метод анализа ДНК применяется и для воссоздания внешности современных людей. Собственно, в судебно-медицинской практике и работает ведущий автор нынешней работы.
Недавно мы писали о том, что анализ древней ДНК помог учёным установить происхождение легендарных этрусков. Рассказывали мы и о том, что по ДНК учёные восстановили внешность денисовского человека, а также о том, что показал анализ ДНК древнейшего жителя Великобритании.
Больше новостей из мира науки вы найдёте в разделе «Наука» на медиаплатформе «Смотрим».
Как определить свой типаж с учетом природных данных / AdMe
Хотя сегодня индустрия красоты изобилует вариантами одежды и косметики на любой вкус и кошелек, очень трудно подобрать то, что действительно подойдет. Американский имиджмейкер еще в 80-е годы выделил 4 женских типажа внешности, на которые стоит ориентироваться при создании индивидуального стиля.
Мы в AdMe.ru решили познакомиться с этой полезной теорией поближе. Тест поможет определить природный типаж и подобрать выигрышные сочетания одежды и макияжа. На каждом этапе вам нужно будет подобрать вариант ответа, который больше соответствует вашей внешности. Расшифровать результат будет проще, если вы будете записывать выбранные варианты.
Костная структура (ширина кости)
Форма плеч
Кисти и стопы
Тип фигуры (мягкие ткани)
Грудь
Талия
Бедра
Руки (от локтя до плеча) и ноги (от колена до бедра)
Подбородок
Нос
Скулы
Форма глаз
Форма губ
Щеки
Волосы
Большинство ответов
А — драматик
Типичный представитель: Тильда Суинтон.
Носить: плотные текстуры с матовой и ровной поверхностью, ткани, которые хорошо держат форму. Следует избегать прозрачных и легких тканей и грубых текстур, например тяжелой вязки.
Макияж: яркие губы и подчеркнутые скулы.
Большинство ответов
Б— натурал
Типичный представитель: Дженнифер Энистон.
Носить: расслабленные и прямые силуэты, можно сочетать между собой цвета, текстуры и узоры в рамках одного образа. Следует избегать монохромных образов, одежды oversize, а также тонких и прозрачных тканей.
Большинство ответов
В
— классик
Типичный представитель: Мерил Стрип.
Носить: гладкие и мягкие силуэты, симметричные и сдержанные узоры и принты. Рекомендуется использовать одинаковые формы в рамках одного образа: или геометричные, или округлые. Следует избегать бесформенных силуэтов и сложных узоров с множеством деталей.
Макияж: матовый.
Большинство ответов
Г— гамин
Типичный представитель: Одри Тоту.
Носить: острые и прямые силуэты, живые принты, подойдет одежда с обилием мелких деталей. Следует избегать длинной вертикальной линии, симметричных силуэтов и грубых текстур.
Макияж: smoky eyes.
Большинство ответов
Д — романтик
Типичный представитель: Скарлетт Йоханссон.
Носить: легкие ткани, которые легко драпируются, замысловатые плавные принты, округлые формы. Следует избегать строгих силуэтов, прямых линий и острых углов.
Макияж: в золотистых, розовых, персиковых тонах.
А вы нашли себя среди типажей? Напишите в комментариях.
Систему предсказания цвета глаз и волос по ДНК уточнили
Работа выполнена в рамках проекта Союзного государства «ДНК-идентификация». Расположение исследованных популяций на карте. 1 — чуваши, 2 — коми-пермяки, 3 — коми-зыряне, 4 — марийцы равнинные, 5 — марийцы горные, 6 — мордва эрзя, 7 — мордва мокша, 8 — русские, 9 — русские (казаки-некрасовцы), 10 — русские из Нижегородской области, 11 — русские из Тверской области, 12 — русские из Ярославской области, 13 — удмурты, 14 — татары Поволжья, 15 — адыгейцы, 16 — аварцы, 17 — азербайджанцы, 18 — даргинцы, 19 — кабардинцы, 20 — карачаевцы, 21 — кумыки, 22 — лезгины, 23 — осетины, 24 — рутульцы, 25 — талыши, 26 — цахуры, 27 — турки-месхетинцы, 28 — башкиры, 29 — лесные ненцы, 30 — ханты, 31 — манси, 32 — шорцы, 33 — сибирские татары, 34 — буряты, 35 — чукчи, 36 — дунгане, 37 — эвенки Дальнего Востока, 38 — эвенки Камчатки, 39 — эвенки Охотского побережья, 40 — казахи, 41 — киргизы, 42 — коряки, 43 — нанайцы, 44 — таджики, 45 — туркмены, 46 — уйгуры, 47 — узбеки, 48 — якуты Дальнего Востока. На карте голубой цвет кружков соответствует Европейской части России; зелёный — Кавказу; фиолетовый — Западной Сибири; красный — Северной и Центральной Азии.
Рисунок из статьи: Balanovska E., Lukianova E., et al.// BMC Genomics 2020 21(Suppl 7):527.
‹
›
Можно ли по ДНК, оставленной на месте преступления, узнать внешность преступника? Сегодня криминалисты могут с большой вероятностью определить по ДНК черты внешности, связанные с пигментацией, то есть цвет глаз, волос и кожи. Для этого нужно провести анализ некоторых изменчивых участков в геноме, отличающихся одним нуклеотидом (одной «буквой» ДНК) — они называются SNP маркерами (от single nucleotide polymorphism). Такой анализ проводят не только криминалисты, но и палеогенетики, исследующие древнюю ДНК, благодаря ему они могут сделать предположения о внешнем облике наших далёких предков.
Генетики создали панель SNP маркеров для предсказания цвета глаз, волос и кожи — HIrisPlex-S. Панель представляет собой набор зондов — коротких последовательностей ДНК (олигонуклеотидов), которые комплементарно связываются с изменчивыми участками в геноме (SNP). Каждый такой зонд снабжён флуоресцентной меткой, так что наличие реакции определяется по свечению. 24 SNP маркера позволяют предсказывать цвет глаз и волос, 17 SNP маркеров — цвет кожи (далее пойдёт речь только о цвете глаз и волос). Панель HIrisPlex-S была разработана на базе западноевропейских геномов (в основном геномов голландцев). И потому она с хорошей точностью работает, предсказывая цвет глаз и волос по ДНК для людей европейского происхождения. Но у людей иного происхождения генетические варианты, связанные с пигментацией волос и глаз, могут быть другими, поскольку в разных популяциях они порой возникали независимо. Проблема повышения точности европейской панели SNP маркеров особенно актуальна для России, где проживает более сотни разных народов.
Для большей части неевропейского населения Северной Евразии характерны тёмные глаза и волосы. Но есть регионы (Сибирь, Урал, Кавказ), где встречается фенотип* со светлыми волосами и светлыми глазами, причём — у коренных народов, которые не состоят в близком родстве с населением Западной Европы. Вполне возможно, что светлый фенотип обеспечивается у них другими генетическими вариантами, чем у западноевропейцев.
И эти генетические варианты надо выяснить, чтобы система предсказания внешности по ДНК могла точно работать в североевразийских популяциях.
Российские генетики и антропологи, статья которых опубликована в журнале «ВМС Genomics», нашли эти новые генетические варианты. В коллектив соавторов вошли специалисты из Института общей генетики РАН, Медико-генетического научного центра, МГУ им. М. В. Ломоносова и Института этнологии и антропологии РАН при участии коллег из других научных организаций в Москве, Краснодаре и Нурсултане (Казахстан). Ведущий автор статьи — доктор биологических наук, профессор Елена Балановская, заведующая лабораторией МГНЦ.
Генетики проанализировали образцы ДНК трёхсот человек из 48 популяций, принадлежащих к коренным народам России и сопредельных стран, эти образцы они взяли из коллекции Биобанка Северной Евразии. Анализ ДНК проводили путём секвенирования экзома (части ДНК, кодирующей белки). Для этих трёхсот человек в коллекции хранились также антропологические фотографии, с которыми работали три независимых эксперта-антрополога.
В итоге была собрана база данных, включившая как генотипы (генетические варианты), так и фенотипы (цвет волос и глаз).
Сначала на этой базе данных протестировали работу системы HIrisPlex-S, и выяснилось, что точность предсказания цвета глаз и волос в этих 48 популяциях несколько ниже, чем в популяциях Западной Европы. Снижение точности особенно заметно для предсказания цвета глаз у представителей народов Кавказа.
На следующем этапе работы исследователи стали искать новые генетические варианты, которые могут быть связаны с пигментацией глаз и волос в популяциях Северной Евразии. Для этого они изучили 117 012 новых SNP в 53 генах и межгенных регионах (участках ДНК, которые не кодируют белки). Эти 53 гена были специально отобраны — те гены, которые вовлечены в генетический контроль пигментации. Используя определённые статистические методы, авторы выявляли генетические варианты, коррелирующие с внешностью, и постепенно сужали этот список, оставляя из них лишь важнейшие. Итоговый список для предсказания цвета глаз включает 36 SNP, из них семь проявляют наибольшую предсказательную силу. Два SNP из этих семи были уже известны и входили в систему HIrisPlex-S, остальные пять ранее не были описаны. Четыре SNP маркера из пяти лежат в известном гене HERC2, связанном с изменчивостью цвета кожи, волос и глаз; пятый SNP расположен в межгенном регионе. Такой же анализ провели для генетических вариантов, определяющих цвет волос. Итоговый список включил 33 SNP, из которых 11 обладали наибольшей предсказательной силой. Три из них были уже известны и включены в панель HIrisPlex-S, остальные восемь ранее не описаны.
«Мы задались целью проверить, будет ли европейская система предсказания цвета глаз и волос по генотипу работать на российских популяциях, — рассказал руководитель исследования Олег Балановский, — ведь многие из них обладают совершенно иным генофондом. Выяснилось, что система работает в целом неплохо, но её можно улучшить, введя в неё новые маркеры, выявленные нами в популяциях Кавказа и Сибири».
Российские генетики считают, что точность предсказания внешности человека по ДНК в нашей стране станет выше при добавлении в стандартную панель HIrisPlex-S этих новых маркеров.
Комментарии к статье
* Фенотип — совокупность внешних и внутренних признаков организма, которая формируется на основе генотипа.
Распознать сексуальную ориентацию человека на основе внешних признаков невозможно
Стереотипы о том, как должны выглядеть и вести себя геи, не соответствуют действительности, выяснили ученые. Но даже те люди, которые называют себя лояльными по отношению к сексуальным меньшинствам, поддаются влиянию стереотипов — до такой степени, что готовы бить геев током.
В психологии существует термин «гейдар» — производное от слов «гей» и «радар». Этим понятием обозначается способность человека определять сексуальную ориентацию других людей исключительно на основе внешних признаков: внешности, манеры общения, роду занятий.
Чаще всего подобные суждения спонтанны и основываются на первом мимолетном впечатлении от собеседника, опираются на стереотипные признаки «мужественности» или «женственности».
Несмотря на это, существование «гейдара» стало объектом исследования психологов, сексологов и антропологов еще в конце ХХ века. Научные статьи о том, что «встроенный радар» для вычисления людей с нетрадиционной сексуальной ориентацией действительно существует, публиковались в достаточно серьезных журналах, например, в The Journal of Personality and Social Psychology (импакт-фактор этого издания равен 5,031) или в чуть менее престижном Archives of Sexual Behavior (импакт-фактор равен 2,589).
The Journal of Personality and Social Psychology еще в 1999 году опубликовал исследование группы ученых, возглавляемой социальным психологом Налини Амбади из Стэнфордского университета. Психологи пришли к выводу, что люди действительно могут определять сексуальную ориентацию человека с большей вероятностью, чем простая случайность, причем достаточно информации для этого могут дать как видеозаписи, так и статичные изображения человека.
Американские ученые под руководством Уильяма Кокса из Висконсинского университета в Мэдисоне провели еще одно исследование феномена «гейдара» и заявили: на самом деле такого явления не существует, а все «признаки гея» — гендерная неконформность, внешность, манера речи — являются не более чем стереотипами. Тем не менее люди (даже те, кто уверяет, что относится к геям абсолютно лояльно) подвержены этим стереотипам до такой степени, что готовы бить геев током. Статьи были опубликованы в журналах Psychological Science и The Journal of Sex Research.
В ходе эксперимента людям демонстрировались фотографии мужчин — как традиционной, так и нетрадиционной сексуальной ориентации. В первом опыте с фотографий были убраны прически мужчин — как утверждают авторы работы, прическу всегда можно легко изменить, и поэтому ее нельзя рассматривать как часть фенотипа человека. Во второй раз снимки мужчин демонстрировались уже целиком. Каждое фото сопровождалось серией подписей-стереотипов, которые изображенные на снимках мужчины делали сами, и рассказывали они о себе при этом правду.
Испытуемым предлагалось ознакомиться с фото и кратким рассказом мужчины о себе и сделать вывод о его сексуальной ориентации. Результаты работы показали: при вынесении решения люди ориентировались не на фенотип человека (так как наличие или отсутствие прически на фото не влияло на «приговор» и вызывало изменение решения лишь в 4–7% случаев), а на подписи — то есть на стереотипы, — которые вызывали 24–28% вариаций, но при этом на самом деле действительность не отражали.
Как выяснилось, геев люди «вычисляют» на основе профессии мужчины (наиболее подозрительными стали парикмахеры, дизайнеры интерьеров и медицинские братья), его внешности (признаками гея считаются хорошая стрижка и модная одежда), предпочтений (тем, кто не хочет быть гомосексуалом в глазах окружающих, не стоит признаваться в любви к походам на мюзиклы и в магазины одежды, к танцам, а также в том, что ему нравится творчество певицы Шер).
Наиболее мужественными и уж точно не геями, по мнению респондентов, являются мужчины, работающие полицейскими или пожарными, небрежно одетые, любящие смотреть спортивные каналы и сами играющие в американский футбол или в баскетбол. Если мужчина при этом водит фургон и поддерживает Республиканскую партию (эксперимент, напомним, проводился в США), это окончательно снимает с него все подозрения в нетрадиционной ориентации.
Таким образом ученые доказали: феномен «гейдара» не подразумевает под собой обладания никакой особой интуицией и основан исключительно на стереотипах. При этом даже
те, кто уверяет, что относится к мужчинам с нетрадиционной ориентации лояльно, влиянию этих стереотипов подвержены.
Это было доказано в ходе эксперимента: людям, которые называли себя носителями прогрессивных взглядов и говорили, что относятся к геям спокойно, предложили сыграть в игру с соперником-мужчиной, о котором сообщалось одно из следующих утверждений: «Он гей», «Он любит ходить по магазинам», «Он не гей», «Он любит ходить по магазинам со своей девушкой».
Очевидно, что второе утверждение косвенно намекает на нетрадицонную ориентацию человека, а последнее – на традиционную. Суть игры заключалась в следующем: когда загоралась красная лампочка, испытуемый должен был нажать кнопку, в результате чего соперника било током. Соперник находился за перегородкой и был невидим, а силу тока участник мог выбирать по своему усмотрению. Целью игры было опередить соперника и нажать кнопку раньше него.
На самом деле соперник был вымышленным и за перегородкой никого не было. Впрочем, сам участник опыта удары током все-таки иногда получал: когда задержка в его реакции превышала определенное время, он испытывал удар той силы, которую до этого адресовал партнеру. В результате работы выяснилось, что
уровень агрессии по отношению к геям был значительно выше: удары тока максимальной силы адресовались им в 20,3% случаев.
В случае же с мужчинами традиционной ориентации этот показатель был равен 14,4%. Вымышленные партнеры, любившие ходить по магазинам одни или с девушкой, получали максимальную «порцию» тока в 18 и 18,5% случаев соответственно.
Авторы исследования призывают всех задуматься о своем отношении к людям нетрадиционной сексуальной ориентации и над тем, на чем основаны их чувства. Кроме того, стало очевидно, что многие из нас — даже если пытаются показаться лояльными — на самом деле относятся к гомосексуалам не так, как к традиционно ориентированным мужчинам и женщинам.
2017 — 2 | Сайт Научно-Практического журнала для врачей «Педиатр»
◆ ПЕРЕДОВАЯ СТАТЬЯ
Г.Ю. Модель, Г.Ф. Коротько
О липолизе в лактотрофии новорожденных и детей первого года жизни
◆ ОРИГИНАЛЬНЫЕ СТАТЬИ
В.С. Василенко, Е.С. Семенова, Ю.Б. Семенова
Липиды крови у спортсменов в зависимости от направленности тренировочного процесса
Д.О. Иванов, Л.В. Козлова, В.В. Деревцов
Вегетативный статус и адаптация у младенцев, имевших разные типы внутриутробной задержки роста
Результаты оценки качества лечения и жизни детей с эпилепсией
И.Б. Ершова, Т.В. Ширина, Т.А. Гончарова
Особенности развития сенсорной функции детей в зависимости от вида вскармливания в условиях военных действий
Т.С. Дьяченко, Л.Н. Грибина, О.Ф. Девляшова
Мнение родителей о работе педиатрических амбулаторно-поликлинических организаций как важный критерий оценки качества медицинской помощи детскому населению
А.В. Миронова, В.Г. Баласанян
Способ оценки и прогнозирования становления менструальной функции у девочек-подростков
Т.В. Брус, М.А. Пахомова, А.Г. Васильев
Коррекция печеночной дисфункции на модели обширного глубокого ожога
◆ ОБЗОРЫ
Е.М. Кучинская, М.М. Костик, В.Г. Часнык
Современные основы формализованного описания течения системной красной волчанки у детей
Неонатальный сахарный диабет в структуре IPEX-синдрома
◆ КЛИНИЧЕСКАЯ ПСИХОЛОГИЯ
Н.В. Козина, М.В. Васильева
Особенности динамики самооценки лиц с расстройствами аутистического спектра в процессе реабилитации
Скачать выпуск 2 2017 года
Материал опубликован 20.05.2017 пользователем Редактор
Олег Балановский выступил с докладом на конференции Международного консорциума по феному человека «2nd Board Meeting of International Human Phenome Consortium»
Вторая конференция Международного консорциума по изучению фенома человека (2nd Board Meeting of International Human Phenome Consortium) в этом году из-за эпидемиологической обстановки проводилась онлайн. Главный научный сотрудник лаборатории популяционной генетики человека ФГБНУ МГНЦ, д.б.н., профессор РАН Олег Балановский выступил с докладом «2D and 3D imaging of the various populations from Russia: updating the pigmentation-predicting system» («2D и 3D фотопортреты различных популяций России: обновление системы предсказания пигментации по ДНК»).
Олег Балановский представил Биобанк Северной Евразии, содержащий 30 тысяч образцов ДНК из 300 популяций коренных народов России и сопредельных стран. Сбор коллекций биологических образцов коренных народов сопровождается антропологической фотографией обследуемых. Суммируя антропологические фотопортреты разных людей, принадлежащих к одной популяции, исследователи получают обобщенный фотопортрет популяции. Антропологические фотопортреты позволяют оценить черты внешности, в частности, связанные с пигментацией, и собрать коллекцию фенотипов коренных народов. Сравнение базы данных по генотипам и фенотипам показало, что система предсказания фенотипа пигментации по ДНК (HIrisPlex-S) в популяциях Северной Евразии работает менее точно, чем в популяциях Западной Европы. Это объясняется тем, что в Северной Евразии могли возникнуть другие аллели, связанные с пигментацией, отличные от западноевропейцев. Коллективом лаборатории популяционной генетики человека были найдены новые SNP маркеры, ассоциированные с цветом глаз и волос в популяциях Северной Евразии. Включение этих маркеров в панель повысит точность предсказания цвета глаз и волос по ДНК в нашей стране. Исследователи ставят перед собой задачу расширить данный подход на другие фенотипические черты. Для этого они используют не 2D фотопортреты, а 3D фотопортреты, получаемые с помощью портативного антропологического сканера.
Сотрудничество в рамках Международного консорциума позволит решить проблему стандартизации антропологических измерений и общими усилиями организовать массовое фенотипическое исследование популяций Китая и России.
Ссылка на доклад https://youtu.be/O4zsyRkpasE
Ссылка на конференцию: https://cwmeet.webex.com/webappng/sites/cwmeet/meeting/download/0ce70f96f8434847b86c5463b0a6844b?MTID=md6fb4126f44c18d9caf33aabbe6d3d0f&siteurl=cwmeet
Эпохальное изменение внешности — ariaryavarta — LiveJournal
«За последние 10-20 тысяч лет резко усилилась инфантильность биологическая. Это выразилось в таких чертах, как: • Округление головы. • Грацилизация челюстей. • Позднее прорезание «зубов мудрости». • Относительное увеличение глаз и уменьшение рта. • Сужение таза. • Более позднее окостенение зон роста. • Облегчение костяка в целом. • Уменьшение контрфорсов и поверхностей для прикрепления мышц (шероховатостей, гребней и др.), что говорит об ослаблении мускулатуры. • Роды в среднем в более позднем возрасте. • Снижение зрелости («доношенности») новорожденных.
В очагах перенаселенности (Индия, Дальний Восток, Европа) усиливаются инфантильные традиции и в культуре. Например: • В очагах перенаселенности мужчины генетически утрачивают бороду. Там, где этого не произошло, хорошим тоном считается брить лицо.
• Развивается нарочито тонкоголосое, слащавое пение, как женщин, так и мужчин (Дальний Восток, Индостан, в ХХ веке – американско-европейская поп-музыка) • Происходит сакрализация образа женоподобного и инфантильного мужчины (крохотный рот, огромные глаза, мягкие манеры). Таковы боги индуизма, китайские и христианские святые. • Меняются тенденции лицедейства. В первобытном обществе на публику работают крупные люди с выступающими, гротескными, пугающими чертами лица. В перенаселенном цивилизованном обществе идеальный актер – женоподобный травести. Там Пьеро побеждает Арлекина и Пульчинеллу.
• В массовой культуре ведущие позиции занимают люди с инфантильной, «феминизированной» внешностью. • Изменяются мораль, этика, право. Еще недавно дети считались наименее ценной прослойкой, а жестокость в их отношении одобрялась, как воспитательная мера и неотъемлемое право взрослых. Теперь всё не так.
• Становится модным молодежный стиль жизни, основная доля которого приходится не на труд и службу, а на обучение, тренировку, игру, посещение брачных ристалищ (дискотек, соревнований, мест активного досуга и других «тусовок»). Люди проводят больше времени в группах с ослабленной иерархической структурой. • Стирается ритуальная грань между детским/подростковым и взрослым стилем оформления – в одежде, аксессуарах (например, рюкзак, канцтовары), интерьере и других атрибутах. • Не берусь утверждать, но полагаю, что подобные тенденции охватывают и язык. Например, в очагах перенаселенности должна проявляться детская «незрелость» артикуляции (нечеткость, вытягивание губ, преобладание высоких тонов, большая амплитуда интонирования и др.). • Эротические каноны смещаются в сторону омоложения. • Люди с инфантильным обликом в среднем получают большую аттрактивность – это модные «красавчики». Особенно в молодежных коллективах (где и осуществляется поиск брачного партнера)».
Генотип против фенотипа: примеры и определения
Любой организм является побочным продуктом как его генетической структуры, так и окружающей среды. Чтобы понять это в деталях, мы должны сначала ознакомиться с некоторыми базовыми генетическими терминами и понятиями. Здесь мы даем определения для терминов генотип и фенотип , обсуждаем их взаимосвязь и смотрим, почему и как мы можем их изучать.
Что такое определение генотипа?
В биологии ген — это участок ДНК, кодирующий признак. Точное расположение нуклеотидов (каждый из которых состоит из фосфатной группы, сахара и основания) в гене может отличаться в разных копиях одного и того же гена. Следовательно, ген может существовать в разных формах в разных организмах. Эти различные формы известны как аллели. Точно фиксированное положение на хромосоме, которое содержит определенный ген, известно как локус.
Диплоидный организм наследует либо две копии одного и того же аллеля, либо одну копию двух разных аллелей от своих родителей. Если человек наследует два идентичных аллеля, говорят, что его генотип гомозиготен по этому локусу.
Однако, если они обладают двумя разными аллелями, их генотип классифицируется как гетерозиготный по данному локусу. Аллели одного и того же гена бывают либо аутосомно-доминантными, либо рецессивными. Аутосомно-доминантный аллель всегда экспрессируется преимущественно по сравнению с рецессивным аллелем.
Последующая комбинация аллелей определенного гена, которой обладает индивидуум, составляет его генотип .
Примеры генотипов
Рассмотрим классический пример – цвет глаз.
Ген кодирует цвет глаз.
В этом примере аллель либо коричневого, либо синего цвета, причем один унаследован от матери, а другой от отца.
Аллель коричневого является доминантным (В), аллель голубого – рецессивным (б). Если ребенок унаследует два разных аллеля (гетерозигота), у него будут карие глаза. Чтобы у ребенка были голубые глаза, он должен быть гомозиготным по аллелю голубых глаз.
Рисунок 1: Диаграмма наследования, показывающая, как человек может наследовать голубые или карие глаза в зависимости от аллелей, переносимых его родителями, при этом аллель карих глаз является доминантной, а аллель голубых глаз — рецессивной.
Другие примеры генотипа включают:
Цвет волос
Рост
Размер обуви
Что такое определение фенотипа? Сумма наблюдаемых характеристик организма составляет его фенотип. Ключевое различие между фенотипом и генотипом заключается в том, что генотип наследуется от родителей организма, а фенотип — нет.
Хотя фенотип влияет на генотип, генотип не равен фенотипу.На фенотип влияют генотип и факторы, включая:
Эпигенетические модификации
Факторы окружающей среды и образа жизни
стать ярко-розовым. Примеры фенотипов Факторы окружающей среды, которые могут влиять на фенотип, включают питание, температуру, влажность и стресс.Фламинго — классический пример того, как окружающая среда влияет на фенотип. Несмотря на то, что они известны своим ярко-розовым цветом, их естественный цвет белый — розовый цвет вызван пигментами в организмах, которые входят в их рацион.
Второй пример — цвет кожи человека. Наши гены контролируют количество и тип меланина, который мы производим, однако воздействие УФ-света в солнечном климате вызывает потемнение существующего меланина и способствует усилению меланогенеза и, следовательно, более темной коже.
Генотип против фенотипа: наблюдение Наблюдать за фенотипом просто – мы смотрим на внешние признаки и характеристики организма и делаем выводы о них.Однако наблюдение за генотипом немного сложнее.
Генотипирование — это процесс, при котором различия в генотипе человека анализируются с помощью биологических тестов. Затем полученные данные можно сравнить либо с последовательностью второго человека, либо с базой данных последовательностей.
Ранее генотипирование позволяло получить только частичные последовательности. Теперь, благодаря крупным технологическим достижениям последних лет, современное секвенирование всего генома.
Рис. 3. Рабочий процесс, изображающий различные этапы полногеномного секвенирования (WGS).
(WGS) позволяет получать целые последовательности. Эффективный процесс, который становится все более доступным, WGS включает в себя использование высокопроизводительных методов секвенирования, таких как секвенирование одной молекулы в реальном времени (SMRT), для определения необработанной последовательности нуклеотидов, составляющих ДНК организма.
WGS — не единственный способ анализа генома организма. Доступны различные методы.
Почему важно изучать генотип и фенотип? Понимание взаимосвязи между генотипом и фенотипом может быть чрезвычайно полезным в различных областях исследований.
Особенно интересной областью является фармакогеномика. Генетические вариации могут возникать в ферментах печени, необходимых для метаболизма лекарств, таких как CYP450. Следовательно, фенотип человека, то есть его способность метаболизировать конкретный препарат, может варьироваться в зависимости от того, какой формой гена, кодирующего фермент, он обладает. Для фармацевтических компаний и врачей эти знания являются ключевыми для определения рекомендуемых дозировок лекарств среди населения.
Использование методов генотипирования и фенотипирования в тандеме, по-видимому, лучше, чем использование только тестов генотипа.В сравнительном клиническом фармакогеномном исследовании мультиплексный подход выявил более значительные различия в способности метаболизма лекарств, чем можно было предсказать только с помощью генотипирования. Это имеет важные последствия для персонализированной медицины и подчеркивает необходимость быть осторожными, полагаясь исключительно на генотипирование.
Как мы можем изучить связь между генотипом и фенотипом? Используя модели животных, таких как мыши, ученые могут генетически модифицировать организм таким образом, чтобы он больше не экспрессировал определенный ген, что известно как нокаутные мыши.Сравнивая фенотип этого животного с фенотипом дикого типа (то есть с фенотипом, существующим, когда ген не удален), мы можем изучить роль определенных генов в появлении определенных фенотипов.
Инициатива Mouse Genome Informatics (MGI) собрала базу данных тысяч фенотипов, которые можно создать и изучить, а также генов, которые необходимо выбить, чтобы получить каждый конкретный фенотип.
Генотип против диаграммы фенотипа:
10
фенотип
Определение
Набор генов в нашей ДНК, который Ответственный за определенную черту
Наблюдаемые характеристики организма
, характеризующиеся
методы генотипирования, такие как WGS
, наблюдение за наружу характеристик организма
зависит от
Генные последовательности, которыми обладает организм
Генотип, ПЛЮС эпигенетика и факторы окружающей среды
Наследуется?
да
№
NO
1010
Гены, кодирующие глазные цвета
человек с коричневыми глазами
5
Определение лица: что такое DNA Thenotyping действительно расскажет нам о Неизвестный образец?
В каждом человеке, где-то среди примерно трех миллиардов пар оснований ДНК, скрытых в аллелях и полиморфизмах одиночных нуклеотидов, находится информация, которая во многом определяет его внешний вид. Этот определяемый ДНК внешний вид, или фенотип, создает семейное сходство и, по словам генетика Ричарда Спритца, «то, на что реагирует ваша бабушка, когда она говорит, что вы похожи на своего отца». Усилия генетиков по поиску фрагментов ДНК, которые определяют, как выглядит человеческое лицо — все, от формы носа до расстояния между глазами, — в последние годы активизировались, и был достигнут прогресс. Теперь ученые могут с некоторой уверенностью использовать цепочку ДНК для определения вероятного цвета волос и глаз человека, а также пигментации кожи и происхождения.Генетик из Пенсильванского государственного университета Марк Шрайвер предпринял то, что он описывает как «первую попытку создания лицевых композитов из ДНК» с «предварительными, но, безусловно, многообещающими результатами».
Эта статья была написана по контракту с NIJ. Первоначально он был опубликован в журнале Forensics Magazine и перепечатывается с разрешения.
Создание «фото» изображения лица человека из нитей ДНК представляет огромный интерес для судебно-медицинских экспертов. Физический портрет подозреваемого может быть разработан на основе ДНК, оставленной на месте происшествия, когда не было свидетелей преступления.После пожаров или других катастроф ДНК неопознанных тел может сделать их узнаваемыми для членов семьи. Кроме того, ДНК из фрагмента кости может помочь визуализировать и идентифицировать людей в массовых захоронениях. Хотя большинство генетиков подчеркивают ограниченность исследований фенотипов, полиция и судебные следователи уже могут обратиться к горстке частных компаний, которые утверждают, что могут использовать ДНК для точного предсказания внешности человека. Итак, каково состояние исследований фенотипа? Может ли судебно-медицинский следователь реально рассчитывать получить точное изображение человека из фрагмента ДНК?
«Мы даже не в конце начала», — сказал Спритц, который шесть лет пытался определить и понять, что определяет внешний вид человеческого лица.Спритц, программный директор Медицинской и генетической программы Университета Колорадо, является одним из нескольких исследователей фенотипов ДНК, получающих поддержку от Национального института юстиции. Хотя генетики с осторожностью относятся к преувеличению прогресса в создании точного физического образа человека на основе ДНК, существует общее мнение, что понимание основ фенотипов значительно продвинулось вперед за последнее десятилетие. Теперь ученые могут использовать ДНК для определения с вероятностью более 75 процентов происхождения человека, цвета глаз и волос.
Большая часть работы по изучению предков выполняется генетиком из Йельского университета Кеннетом Киддом, который разработал панель из 55 «информативных о происхождении одиночных нуклеотидных полиморфизмов» (AISNP), которые делят людей на восемь географических регионов, таких как Европа, Восточная Азия и Тихоокеанский. Например, ДНК из фрагмента кости, найденного во Вьетнаме, можно сравнить с панелью AISNP, чтобы определить, был ли человек из Юго-Восточной Азии или Северной Америки. Однако если бы этот человек был афроамериканцем, результаты оказались бы эфиопскими, потому что это смесь европейских и африканских генов. Кидд расширяет панель AISNP, чтобы включить больше географических регионов. Определение происхождения человека — это часть генетической головоломки, которая определяет, как выглядит этот человек, и это важная часть более широких усилий по точному изображению конкретного лица по ДНК. Несмотря на прогресс, которого он добивается, Кидд сказал, что ему еще предстоит пройти долгий путь. «С такими исследованиями, которыми я занимаюсь, — сказал он, — я никогда не закончу».
Больше, чем просто «коричневый против синего»
В исследовательской лаборатории Университета Индианы и Университета Пердью в Индианаполисе генетик Сьюзен Уолш работает над усовершенствованием фенотипирования ДНК для предсказания количественного цвета — или точного цвета глаз, волос и кожи.Более ранняя работа Уолша и других идентифицировала однонуклеотидные полиморфизмы или SNP, которые вызывают пигментацию. «Это категорическая идентификация, — сказала она, — карие глаза против голубых, светлые волосы против каштановых. Нашей целью сейчас является определение реального цвета, как значение RGB в Adobe Photoshop». Что вдохновило ее на определение реального цвета по ДНК, так это просьба молекулярного генетика Тури Кинга определить цвет глаз и волос Ричарда III, останки которого были найдены под парковкой в Лестере, Англия, в 2012 году.Кинг, который использовал митохондриальную ДНК, чтобы подтвердить, что останки принадлежат Ричарду III, обратился к Уолшу, чтобы определить, какой из портретов короля — все они были написаны после того, как он был убит в бою в 1485 году — был наиболее точным. Основываясь на анализе фенотипа Уолша, Кинг определил, что одна из самых ранних картин Ричарда III, «Портрет в арочной раме» 1510 года, лучше всего соответствует генетической информации.
«Мы все еще имели дело с категориями [цвета], потому что мы еще не достигли количественного уровня», — сказала Уолш о своем определении цвета волос и глаз Ричарда III.«[Кинг] хотел увидеть что-то физическое, и именно это побудило меня так сильно двигаться в сторону количественных показателей. Потому что я всегда мог сказать кому-нибудь «голубой» или «блондин», а они бы сказали: «Мне нужно увидеть это физически». Вот над чем я сейчас работаю. Я хочу добиться такого результата». Уолш собрал данные о фенотипах ДНК от 2000 человек из Ирландии, Греции и США и в настоящее время собирает данные еще от 3000 человек из тех же стран, чтобы создать базу данных фенотип-генотип и модель прогнозирования.Для судебно-медицинских целей она хотела бы иметь возможность начать с «пустого человека» и по образцу ДНК определить настоящую пигментацию глаз, волос и кожи.
В своей лаборатории в Колорадо Спритц использовал «ориентацию», чтобы попытаться идентифицировать и понять сложные взаимодействия генов, которые определяют структуру лица. Он использовал 29 лицевых ориентиров, стандартизированных в этой области, как способ соотнесения структур лица с генами, но в настоящее время он движется к новому подходу. Спритц подозревает, что основные методы, используемые в большей части работы с лицом, не являются «правильным способом думать о форме лица, потому что ни один из них на самом деле не сработал. «Предыдущая работа, связанная с линейным измерением между ориентирами, не сработала. Итак, его новый подход состоит в том, чтобы сосредоточиться на ориентирах, которые имеют самый высокий генетический компонент наследуемости. «Вот с чего мы начнем», — сказал Спритц. «Мы очень сильно скрестили пальцы, чтобы это выглядело лучше».
Spritz отметил, что генетика — это лишь часть того, что определяет внешний вид человеческого лица. «Окружающая среда и случайность играют большую роль», — сказал он, хотя точно неясно, насколько сильно они влияют.Однако «чем меньше роль играют гены, тем меньше у вас будет возможности когда-либо теоретически собрать все воедино, чтобы сделать фотографию того, как выглядит человек».
Шрайвер, споривший со Spritz в журнале PLOS Genetics о возможности предсказания формы и внешнего вида лица, сказал, что понимание каждой генетической детали, биологического шага и механизма не обязательно для предсказания того, как будет выглядеть лицо. «Вам не нужно понимать механизм, чтобы делать прогнозы, чтобы вывести статистическую закономерность», — сказал Шрайвер. «Было бы ошибкой утверждать, что это слишком сложный процесс для нашего понимания. Тело это понимает. Есть все эти сложные взаимодействия, и они считываются телом. Есть ли в этом что-то особенное, что означает, что мы не сможем это понять?»
Шрайвер заметил, что когда люди смотрят друг на друга, они подсознательно считывают гены, которые являются результатом эонов естественного отбора. Лицо «отражает генетические различия между популяциями, и мы думаем, что это потому, что лицо сыграло важную роль в эволюции нашего вида», — сказал он.
«Мы взаимодействуем друг с другом, основываясь на наших лицах, и естественный отбор определяет, считаем ли мы это лицо привлекательным или нет. Мы ищем лица, похожие на нас самих. . . и смотрим на доминантность лица, выглядит ли человек круче меня. Такого рода вещи привели к быстрой эволюции лица, поэтому эти гены претерпели ускоренную эволюцию».
Независимо от направленности и подхода генетиков к исследованию фенотипа ДНК, поиск генетических основ человеческого лица остается сложной задачей. «Есть некоторые генетики, которые относительно просты, например, болезнь», — сказал Спритц. «Есть промежуточные значения, такие как рост человека, а есть невообразимо сложные, такие как определение формы и черт лица».
Криминалистическое ДНК-фенотипирование: предсказание внешности человека по материалам с места преступления для следственных целей предсказание внешности человека по судебно-медицинским образцам.
•
FDP предназначен для проведения полицейских расследований по делам, в которых нет известных подозреваемых.
•
FDP включает судебно-медицинское использование ДНК для расследования, а не в зале суда.
•
На сегодняшний день признаки пигментации человека предсказуемы как цветовые категории по ДНК.
•
FDP дополнительных черт внешности зависит от обширных будущих исследований.
•
Сегодня мы далеки от способности ДНК предсказывать лица людей.
Abstract
Судебно-медицинская экспертиза ДНК Фенотипирование относится к прогнозированию черт внешности неизвестных доноров образцов или неизвестных умерших (пропавших без вести) лиц непосредственно по биологическим материалам, обнаруженным на месте происшествия. Результаты судебного фенотипирования ДНК в качестве «биологических свидетелей» могут предоставить следственные данные для отслеживания неизвестных лиц, которые не могут быть идентифицированы с помощью текущего сравнительного ДНК-профилирования. Это разведывательное применение ДНК знаменует собой существенно иное судебно-медицинское использование генетического материала, а не текущее профилирование ДНК, представленное в зале суда.В настоящее время групповые признаки пигментации уже предсказуемы по ДНК с достаточно высокой точностью, в то время как несколько других внешне видимых характеристик находятся в стадии генетического исследования. До тех пор, пока индивидуальный внешний вид не станет точно предсказуемым по ДНК, обычное профилирование ДНК необходимо выполнять после предсказания внешнего вида ДНК. Примечательно, и там, где криминалистическое фенотипирование ДНК показывает большие перспективы, это относится к (гораздо) меньшей группе потенциальных подозреваемых, которые соответствуют характеристикам внешности, предсказанным ДНК по пятну с места преступления или по останкам умершего человека.Ожидается, что при наличии достаточного финансирования будущие исследования, направленные на лучшее понимание генетической основы внешности человека, приведут к значительно более подробному описанию внешности неизвестного человека на основе ДНК, что повысит ценность полицейских расследований уголовных дел и дел о пропавших без вести с участием неизвестных.
Использование фенотипирования для предсказания внешности
Как и многие девочки-подростки 1990-х годов, доктор Сьюзен Уолш была очарована работой агента Даны Скалли в качестве патологоанатома в телесериале «Секретные материалы». Уже заинтересованный в понимании того, как люди работают и функционируют, наблюдение за тем, как агент Скалли использует науку, чтобы попытаться раскрыть крайне необычные случаи, вдохновило Сьюзен на получение степени в области биохимии в Университетском колледже в Корке, Ирландия, а затем на степень магистра в области судебно-медицинского профилирования ДНК в Университет Центрального Ланкашира.
Затем она защитила докторскую диссертацию по судебной генетике в Университете Эразма и приехала в США, чтобы получить постдокторскую степень в Йельском университете.
«… Когда я был готов сделать докторскую, я долго думал. И это то, что я также говорю студентам: «Не прыгайте сразу в аспирантуру, это от 4 до 5 лет вашей жизни, так что вы должны сдерживаться и действительно, действительно убедиться, что тема, которую вы повторный выбор — это то, что вы хотите сделать. Я буквально только что погуглил «ДНК судебной экспертизы внешности» и наткнулся на Dr.Манфред Кайзер. Я отправил ему электронное письмо, а остальное уже история».
После получения докторской степени в Йельском университете она подала заявку на должность профессора биологии и судебной медицины в Университете Индианы и Университете Пердью в Индианаполисе и работает там с августа 2014 года. полученная в области судебной экспертизы в результате фенотипирования ДНК, она хотела исследовать методологии, которые позволили бы получить больше информации из ДНК, взятой из образцов с места преступления.Более ранняя работа доктора Уолша была сосредоточена на использовании SNP для улучшения понимания генетики, лежащей в основе определения внешнего вида и происхождения человека. Эта информация может быть использована в контексте судебно-медицинской экспертизы для предоставления правоохранительным органам ключевой разведывательной информации, такой как цвет глаз, волос или кожи. Хотя эта информация не является исчерпывающей, она может быть использована для того, чтобы дать указания правоохранительным органам в качестве следственного ориентира в сложных делах о пропавших без вести или в ситуации массового бедствия.
«Поскольку мы имеем дело с количественными признаками, где различные гены, а также факторы окружающей среды могут влиять на фенотип, мы пытаемся раскрыть генетическую основу определенных признаков, используя, среди прочего, общегеномные ассоциативные подходы и секвенирование следующего поколения. Затем мы пытаемся определить, какие биомаркеры являются наиболее предсказуемыми, и разрабатываем молекулярные инструменты для прогнозирования этих признаков для практического применения в криминалистике и антропологических исследованиях».
Границы | Дифференциальный взгляд на отношения генотип-фенотип
Введение
Иногда кажется, что мы забываем, что первоначальный вопрос генетики заключался не в том, что делает белок, а в том, что делает собаку собакой, а человека мужчиной.
(Благородный, 2006)
Один из фундаментальных вопросов биологии состоит в том, чтобы понять, что отличает особей, популяции и виды друг от друга. Концепция фенотипа , который соответствует наблюдаемым признакам индивидуума, была придумана в противовес генотипу , наследственному материалу, передаваемому гаметами. Начиная с раннего предположения, что генотипы и фенотипы образуют два принципиально разных уровня биологической абстракции (Johannsen, 1911), задача состояла в том, чтобы понять, как они взаимодействуют друг с другом, как генотипы отображаются на фенотипы.За последние 15 лет более 1000 примеров изменений последовательности ДНК были связаны с естественными невредными фенотипическими различиями между особями или видами у эукариот (Martin and Orgogozo, 2013b). У человека каталог OMIM ® (Online Mendelian Inheritance in Man, http://omim.org/), в котором собраны генетические детерминанты фенотипов, связанных с заболеванием, насчитывает более 4300 записей и в общей сложности 2493 опубликованных полногеномных ассоциативных исследования. (GWAS) обнаружили множество участков в геноме, которые статистически связаны со сложными признаками (Welter et al., 2014). Поскольку обнаружение причинно-следственных связей между генетическими и фенотипическими вариациями ускоряется, пересмотр наших концептуальных инструментов может помочь нам найти объединяющие принципы в рое данных. Здесь мы размышляем о взаимосвязи между генотипами и фенотипами и адресуем это эссе биологам, которые готовы попытаться бросить вызов своему нынешнему пониманию фенотипов. Выделим одну полезную точку зрения — дифференциальную. Затем мы показываем, что эта простая структура остается полезной в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды.
Гены как создатели различий
Мутации, выделенные из лабораторных штаммов, сыграли важную роль в понимании карты GP. В классической схеме мутацию сравнивают с эталоном дикого типа, и ее фенотипические эффекты используются для вывода о функции гена. Эта структура часто приводит к семантическому сокращению: от генетического изменения, вызывающего 90 318 вариацию 90 319 фенотипа, часто бывает удобно ассимилировать соответствующий ген в качестве причинной детерминанты признака (Keller, 2010; рисунок 1A).Часто можно встретить заголовки, выражающие эти упрощения, трубящие перед широкой аудиторией об открытии гена «долголетия» или «благополучия», которые приносят научную точность в жертву психологическому воздействию. В соответствии с этим следует ли называть ген, мутация которого летальна, «геном жизни»? На самом деле эти чрезмерно упрощенные формулировки означают, что 90 318 вариаций 90 319 в данном гене вызывают 90 318 вариаций 90 319 в данном фенотипе (Dawkins, 1982; Schwartz, 2000; Waters, 2007). На самом деле ген сам по себе не может ни вызвать наблюдаемый фенотипический признак, ни быть необходимым и достаточным для появления наблюдаемых признаков.Генам нужна клеточная среда, совместное действие множества других генов, а также определенные физико-химические условия, чтобы оказывать заметное влияние на организмы (рис. 1В). Например, пигментация каштановых волос у одного человека является результатом не только действия генов, кодирующих ферменты, синтезирующие пигмент, но и наличия клеток, продуцирующих пигменты соответствующих молекул субстрата (таких как тирозин для меланина), и количества полученного вещества. солнечный свет (Liu et al., 2013). Таким образом, генетический редукционистский подход, который исследует лишь несколько генетических параметров среди множества причинных факторов, бесполезен для полного решения широкого вопроса о том, что делает волосы коричневыми, что порождает определенную биологическую структуру или процесс в целом.Тем не менее, генетический редукционизм может быть совершенно подходящим для идентификации генетических локусов, где изменение вызывает фенотипические различия (рис. 1С). Различие в цвете волос между двумя людьми в некоторых случаях может быть связано с их генетическими различиями. Заметим, однако, что не все фенотипические изменения можно отнести к генетическим изменениям. Различия в цвете волос также могут быть вызваны негенетическими факторами, такими как возраст, интенсивность солнечного излучения или окраска волос, или сочетанием как генетических, так и негенетических различий.
РИСУНОК 1. Схематическое изображение каналов GP. (A) Традиционное представление в классическом генетическом редукционизме. (B) Интегративный взгляд на биологию развития. (C) Схема экспериментального подхода в генетике. (D) Схема экспериментального подхода в эволюционной генетике. (E) Один из примеров таксономически надежного родства GP: SLC45A2 и внутривидовые различия в пигментации у тигров и кур.Отношения GP обозначены пунктирными линиями на панелях (D-E) .
В то время как современная генетика находилась в зачаточном состоянии, Альфред Стертевант сформулировал вопрос о карте ВП простыми словами: «Одной из центральных проблем биологии является проблема дифференцировки — как яйцо развивается в сложный многоклеточный организм? Это, конечно, традиционная главная проблема эмбриологии; но он также появляется в генетике в форме вопроса: как гены производят свои эффекты?» (Стёртевант, 1932). Долгое время некоторые генетики могли думать, что они анализируют морфогенетические механизмы, лежащие в основе формирования фенотипических признаков, в то время как их экспериментальный подход фактически выявлял гены, отсутствие или изменение которых (мутации, делеции, дупликации, перестройки и т. д.) приводит к фенотипические различия (сравните рисунок 1А с рисунком 1С). На самом деле предложение «ваши волосы каштановые» можно интерпретировать либо как абсолютное наблюдение (описание определенного набора молекул, содержащих определенные уровни темного пигмента эумеланина и светлого пигмента феомеланина), либо как имплицитную ссылку на другие возможности. (он коричневый, а не другого цвета).Заблуждения возникают из-за того, что фенотипы обычно определяются относительно возможностей, которые не сформулированы явно. Наше сознание и наш язык часто склонны путать объекты, изменчивость которых рассматривается, с самой изменчивостью (Keller, 2010), и важно напомнить, что в генетике интересующие нас объекты (например, данный генотип, аллель или фенотип) заслуживают определения 90 318 относительно 90 319 по отношению к другому эталонному состоянию.
Таким образом, классический генетический редукционистский подход по своей сути неспособен объяснить все факторы, ответственные за наблюдаемые характеристики в живом мире (Stotz, 2012), но является мощным и актуальным методом анализа генетических рычагов наследуемой фенотипической изменчивости.Сосредоточение внимания на фенотипической изменчивости между особями, а не на абсолютных признаках, присутствующих в отдельных организмах, является ключом к лучшему пониманию генетических причин фенотипического разнообразия.
Связь ГП находится между двумя уровнями вариации
Мышление с точки зрения различий делает очевидным абстрактную сущность, которая заключает в себе как генетический, так и фенотипический уровни. Этот объект состоит из вариации генетического локуса (два аллеля), связанного с ним фенотипического изменения (два различных фенотипических состояния) и их взаимоотношений (рис. 1D).Мы втроем называем совокупность этих элементов «гефе», но здесь мы называем это просто «отношением генотип-фенотип» (отношением ГП). Мы покажем, что отношения ГП — это гораздо больше, чем простое и слабо определенное взаимодействие между двумя уровнями организации: это причинно-следственная связь, которая облегчает наше понимание фенотипического разнообразия.
Генетическая часть отношений с врачами общей практики
В современных базах данных аннотаций генома ген обычно определяется как фрагмент нуклеиновых кислот, который транскрибируется и кодирует РНК или полипептид с известной или предполагаемой функцией (Gerstein et al., 2007). Генетический локус, лежащий в основе фенотипических различий, не обязательно является геном в строгом смысле; он может охватывать конкретную пару оснований, кодирующую область, цис- регуляторную область или распространяться на весь ген с его цис -регуляторными областями или даже на генный кластер (таблица 1). Как ранее отмечалось другими (Falk, 1984; Gilbert, 2000; Stern, 2000; Moss, 2003; Griffiths and Stotz, 2013), концепция гена в биологии развития и в современных базах данных аннотаций генома отличается от концепции гена в эволюционная биология. Здесь акцент делается не на сам ген, как он определен в геномных базах данных, а скорее на индивидуальном функциональном разделении генома на определяющие различия локусы. Генотипическая часть родства GP может принимать форму различных аллелей: разные кодоны, кодирующие разные аминокислоты, вставки/делеции в последовательности, кодирующей белок, расходящиеся версии конкретного регуляторного элемента cis-, наличие/отсутствие вставок транспозонов. , количество копий генов в кластере генов, склонных к структурной изменчивости, и т. д.В геноме не все нуклеотидные участки связаны с фенотипической изменчивостью. Например, вероятно, существуют фрагменты нуклеотидных последовательностей, в том числе так называемая мусорная ДНК (Graur et al., 2015), присутствие которых не оказывает никакого влияния на наблюдаемые характеристики организма, кроме того, что они реплицируются и, возможно, транскрибируются. Существуют также генетические локусы, которые могли быть связаны с фенотипической изменчивостью в прошлом и больше не связаны с фенотипической изменчивостью. Например, генетическая изменчивость в кодирующих областях, связывающих гистоновую ДНК, могла быть важной во время ранней эволюции эукариотических клеток, но эти генетические локусы больше не содержат фенотипически релевантных вариаций, кроме летальных мутаций. Таким образом, в геноме есть участки нуклеотидов, которые абсолютно необходимы для жизни, но сами по себе не содержат жизнеспособных фенотипически релевантных вариаций.
ТАБЛИЦА 1. Несколько примеров отношений GP.
Фенотипическая часть отношений GP
Фенотипический аналог отношения GP относится к разновидности вариации (цвет волос, уровень устойчивости к токсинам и т. д.).), а не к состоянию (светлые волосы, дегустация фенилтиокарбамида и др.; табл. 1).
Фенотип, связанный с генетическим изменением, не обязательно ограничивается организмом, содержащим генетическую мутацию. Например, различие между лево- и правозакрученными раковинами у улитки Lymnaea peregra определяется одним генетическим локусом с материнским эффектом: генотип матери, а не самой особи, отвечает за направление скручивание оболочки (Boycott et al. , 1931). В других случаях причинное генетическое изменение связано с симбионтными бактериями: температурная толерантность тли может различаться у разных особей из-за точечной мутации в их бактериальном симбионте (Dunbar et al., 2007). Определенные фенотипические эффекты могут проявляться и на более высоком уровне, чем организм, несущий генетическое изменение (Dawkins, 1982), например, социальная организация муравьиной колонии (Wang et al., 2013).
Дифференциальная часть отношений общей практики
Как определено выше, родство GP включает в себя генетическое различие и фенотипическое различие.Связь различия как на генетическом, так и на фенотипическом уровне весьма абстрактна и может соответствовать трем различным различиям в живом мире: (#1) различие между двумя репродуктивно изолированными таксонами (живущими или вымершими), (# 2) различие, сегрегирующее внутри популяции, и (3) различие, впервые появившееся в ходе эволюции, между организмом, имеющим наследственный аллель/признак, и его прямым потомком, развившим новый аллель/признак. Следует отметить, что изменчивость фенотипа не всегда сразу следует за появлением новой вызывающей мутации, но может появиться позже из-за единичного набора аллелей, сегрегирующих в популяции.Например, новый фенотип редуцированных панцирных пластин появился в популяции пресноводной колюшки, когда рецессивный аллель EDA , уже присутствующий на криптических уровнях, оказался в гомозиготном состоянии у одной особи (Colosimo et al., 2005; Jones et al., 2012). Главный концептуальный прорыв, сделанный Чарльзом Дарвином, заключался в том, чтобы связать изменчивость среди особей внутри скрещивающейся группы (различие № 2) с изменчивостью между таксономическими группами в пространстве и времени (различие № 1; Левонтин, 1974а).
Отметим также, что определенные фенотипические изменения могут проявляться на уровне всего организма, когда «причинная» мутация сопровождается дополнительными соматическими мутациями, которые весьма вероятны. Например, у женщин, несущих аллель дикого типа и мутантный аллель BRCA1 , клетки могут продуцировать белки BRCA1 дикого типа, поскольку они несут одну копию аллеля BRCA1 дикого типа. Тем не менее, у этих женщин риск развития рака груди или яичников к 70 годам составляет до 80% по сравнению с женщинами, имеющими две копии дикого типа BRCA1 , из-за появления дополнительных вредных мутаций в диком типе BRCA1 . аллель в их соматических клетках молочной железы (Narod and Foulkes, 2004).
Важно отметить, что разница в GP всегда определяется по отношению к интересующей популяции или таксону (Sober, 1988). В менее развитых с медицинской точки зрения странах люди, несущие две дефектные копии гена фенилаланингидроксилазы, имеют серьезные проблемы со здоровьем, включая судороги и умственную отсталость. Напротив, в большинстве развитых стран такие люди диагностируются при рождении и имеют нормальную продолжительность жизни с нормальным умственным развитием благодаря диете с ограничением фенилаланина (Armstrong and Tyler, 1955).Таким образом, взаимосвязь с GP, включающая мутацию с дефектом фенилаланингидроксилазы, зависит от контекста: мутация связана с проблемами со здоровьем в менее развитых в медицинском отношении странах, но не в других странах. Этот пример показывает, что причинно-следственная связь между генетическим изменением и связанным с ним фенотипическим изменением может скрывать множество встроенных параметров (таких как медицинская практика в случае фенилаланингидроксилазы) в рамках предположения при прочих равных «при прочих равных условиях.
Таким образом, связь GP лучше всего рассматривать как связь между двумя вариациями, одна на уровне генотипа, а другая на уровне фенотипа. Человеческий разум может разрабатывать понятия возрастающей абстракции: понятия вещей (например, клетки), понятия изменения (например, эволюции) и понятия отношений (например, гомологии; Cassirer, 1910; Simondon, 1968). Здесь концепция родства ГП устанавливает связь между двумя изменениями (генетическими и фенотипическими). В следующих абзацах мы покажем, что по сравнению с использованием интуитивных представлений о вещах этот обходной путь через усиление абстракции может оказаться более эффективным для лучшего понимания фенотипического разнообразия.
Несколько текущих представлений о связи между генотипом и фенотипом неявно отвергают дифференциальный взгляд
Выше мы утверждали, что при размышлениях о связи между генотипами и фенотипами всегда следует помнить о дифференциальном подходе. GWAS, представляющие собой наиболее популярный метод выявления геномных локусов, ассоциированных со сложными признаками в популяциях, основаны на анализе различий (Visscher et al., 2012). Тем не менее, в современных исследованиях дифференцированная точка зрения иногда неявно отвергается.Когда наблюдается влияние нескольких факторов на фенотипические черты (рис. 1B), дифференциальный взгляд считается слишком упрощенным, и исследователи часто предпочитают снова сосредоточиться на фенотипах отдельных особей, не связывая их явно с фенотипической ссылкой.
В большинстве современных статей проблема связи генотипа с фенотипом формулируется в терминах карт генотипа и фенотипа. Первая карта ВП была представлена Ричардом Левонтином в его книге «Генетическая основа эволюционных изменений» (Левонтин, 1974а; рис. 2А).Он указал средний генотип популяции как точку в пространстве всех возможных генотипов (пространство G) и средний фенотип той же популяции как соответствующую точку в пространстве всех возможных фенотипов (пространство P). Таким образом, эволюционный процесс был разбит на четыре этапа: (1) средний фенотип возникает в результате развития различных генотипов в различных условиях; (2) миграция, спаривание и естественный отбор действуют в пространстве P, чтобы изменить средний фенотип исходной популяции на средний фенотип особей, которые будут иметь потомство; (3) идентичность успешных родителей определяет, какие генотипы сохраняются; и (4) генетические процессы, такие как мутация и рекомбинация, изменяют положение в G-пространстве.
В другом распространенном графическом представлении (рис. 2B) точка в пространстве G и соответствующая ей точка в пространстве P соответствуют генотипу и фенотипу отдельной особи (Fontana, 2002; Landry and Rifkin, 2012). При таком представлении абстрактный объект, который мы определили выше как отношение ГП, будет соответствовать «ходу» в пространстве генотипов, связанному с «ходом» в пространстве фенотипа (или, лучше, сумме нескольких «ходов» в генотипе, и пространства фенотипов, потому что несколько различных геномов могут нести два альтернативных аллеля данного родства GP). В третьем представлении, предложенном Вагнером (1996; рис. 2С), отдельные гены связаны с отдельными признаками.
Хотя эти три графических изображения карт ВП могут облегчить наше понимание некоторых аспектов биологии, во всех из них взаимосвязь ВП и дифференциальное представление понять нелегко. Вызывает недоумение тот факт, что первым человеком, нарисовавшим такую карту ВП, был Ричард Левонтин, красноречивый сторонник дифференциального взгляда (см., например, его предисловие к Oyama, 2000, шедевр убеждения).Поскольку эти графики сосредоточены на индивидуальных, а не на дифференциальных объектах, мы полагаем, что эти три репрезентации имплицитно побуждают нас вернуться к более интуитивной идее об одном генотипе, связанном с одним фенотипом. Упускание из виду дифференциального взгляда также может происходить с точки зрения молекулярной биологии, где белки рассматриваются как имеющие причинные эффекты сами по себе, такие как фосфорилирование субстрата или связывание с последовательностью ДНК. Из-за двух запутанных определений гена, либо как кодирующего белок, либо как вызывающего фенотипическое изменение (Griffiths and Stotz, 2013), легко перейти от дифференциального взгляда к недифференциальному взгляду на отношения GP.
Таким образом, многие современные ментальные представления о связи между генотипом и фенотипом имплицитно отвергают дифференциальную точку зрения. Теперь мы покажем, что дифференциальная точка зрения совместима с тем фактом, что на фенотипические черты влияет сложная комбинация множества факторов, и что мы можем найти соответствующее схематическое представление взаимосвязей GP.
Проблема плейотропии
Разложение организма на элементарные единицы, такие как анатомические структуры, сыграло важную роль во многих биологических дисциплинах, таких как физиология, палеонтология и эволюция.Однако вопрос состоит в том, чтобы определить разложение на признаки, наиболее адекватное интересующему вопросу. Для вопросов, связанных с отношениями между органами различных особей или видов (таких как гомология), может быть уместно сохранить традиционную декомпозицию на анатомические структуры (Wagner, 2014). Рихард Левонтин и Гюнтер Вагнер определили характеры как элементы внутри организма, которые отвечают на адаптивные вызовы и представляют собой квазинезависимые единицы эволюционных изменений (Левонтин, 1978; Вагнер, 2000).Их определение касается абсолютных признаков, наблюдаемых у отдельных организмов (например, форма крыла или количество пальцев у особи), и поэтому далеко от дифференциального взгляда. Здесь, чтобы лучше понять эволюцию и фенотипическое разнообразие живого мира, мы предлагаем разложить наблюдаемые признаки организма на множество элементарных вариаций ГП, которые накапливались в течение многих поколений, начиная с исходного состояния. Мы настаиваем на том, что с этой точки зрения персонажи являются не конкретными объектами (например, кожей), а абстрактными объектами, определяемыми существованием различий между двумя возможными наблюдаемыми состояниями (например, цветом кожи).В качестве аналогии можно представить два способа изготовления изношенной кожаной обуви определенной формы. Можно либо собрать разные атомы в одну организацию, либо можно купить в магазине ботинок, а затем подвергнуть его ряду механических воздействий. Мы естественно склонны сравнивать организмы с машинами и мыслить в терминах частей, которые должны быть собраны, чтобы создать функциональное целое. Однако безудержной метафоры дизайнера или создателя недостаточно для понимания происхождения современных организмов (Coen, 2012).Чтобы понять фенотипические особенности данного организма, более эффективно разложить его на абстрактные изменения, которые происходили последовательно на протяжении эволюционного времени, а не на протяжении времени развития. Исходное состояние является гипотетическим предком изучаемого организма.
Наблюдается, что некоторые мутации (квалифицированные как плейотропные) поражают сразу несколько органов, в то время как другие изменяют только один за раз (Paaby, Rockman, 2013; Zhang, Wagner, 2013). Для плейотропных мутаций мы считаем, что связь ГП должна включать все фенотипические изменения (в разных органах, на разных стадиях и т. ), связанные с генетической разницей. Например, мутация V370A рецептора EDAR связана не только с толщиной волос, но и с изменением плотности потовых и молочных желез в азиатских популяциях (Камберов и др., 2013). Отношения GP в таких случаях являются один-ко-многим. Рассмотрение кожи и глаза как независимых анатомических модулей человеческого тела может показаться уместным для многих эволюционных изменений, но несколько неадекватным в тех случаях, когда эти два органа развили новый признак пигментации одновременно посредством единственной мутации в гене SLC45A2 (Liu и другие., 2013). Рассуждения в терминах взаимосвязей ГП снимают проблему нахождения соответствующей декомпозиции на элементарные анатомические структуры. Сами элементарные отношения ГП представляются адекватными полунезависимыми модулями, комбинация которых может объяснить наблюдаемые характеристики организма.
Проблема непрерывных комплексных признаков
В соответствии с дифференциальной концепцией отношений ГП одним из важнейших моментов является разложение наблюдаемых признаков на ряд полунезависимых фенотипических вариаций, то есть выявление элементарных изменений, происшедших в ходе эволюции. Доступны экспериментальные подходы для разложения данного фенотипического различия на соответствующие более тонкие подвариации. Например, скрещивание растений с разной формой листьев дает потомство, которое демонстрирует составной набор промежуточных форм листьев. Анализ основных компонентов выявил элементарные изменения формы листа, которые вместе могут объяснить разницу в форме между родительскими линиями и, по-видимому, вызваны различными геномными областями (Langlade et al., 2005). Это в какой-то степени предполагает, что «сумма затемняет части.«То, что мы традиционно считаем сложными чертами, можно составить из более простых черт, более поддающихся генетическому анализу. Другим ярким примером является пигментация брюшка в группе Drosophila dunni . Взятые как одна переменная, уровни пигментации демонстрируют сложную генетическую архитектуру, но разложение взрослых паттернов на анатомические субъединицы раскрывает дискретный генетический контроль для каждого субпризнака (Hollocher et al. , 2000). Более известным случаем является эволюция цвета тела пляжных мышей.Различие в окраске между светлыми пляжными мышами и темными мышами можно разложить на отдельные фенотипы (оттенок спины, яркость спины, ширина полосы на хвосте и дорсовентральная граница), которые все связаны с различными мутациями в гене Agouti . Linnen et al., 2013; рис. 3). Каждый генетический локус Agouti , по-видимому, предназначен для спецификации пигментации в данной части тела. Вместе они образуют группу тесно связанных локусов, связанных с изменениями пигментации шерсти.
РИСУНОК 3. Эволюция светлоокрашенных пляжных мышей вызвана несколькими мутациями с отчетливыми эффектами пигментации в локусе Agouti . Темный и светлый фенотипы можно разложить на четыре фенотипических признака, которые связаны с различными однонуклеотидными полиморфизмами (SNP, цветные точки), расположенными в гене Agouti . Показаны только SNP с предполагаемым коэффициентом отбора светлого аллеля выше 0,1. Кодирующие экзоны представлены темными прямоугольниками, а нетранслируемые экзоны — белыми прямоугольниками.Адаптировано из Linnen et al. (2013).
Хотя сложные признаки не всегда можно свести к набору простых взаимосвязей ГП, возможно, что такие признаки, как рост взрослого человека, наиболее показательный количественный признак, который, по прогнозам, состоит из множества генетических эффектов небольшого размера (Fisher, 1930), могут также разложить на элементарные вариации, каждая из которых объясняет более дискретные субпризнаки. В то время как некоторые детерминанты роста человека, такие как LIN28B , были связаны с ростом взрослого человека в разном возрасте, другие гены достигли статистической значимости только в стадийных исследованиях, посвященных росту плода и скорости роста в период полового созревания (Lettre, 2011).Другими словами, эти данные предполагают, что рост человека может быть составным признаком, который модулируется несколькими отношениями GP, каждое из которых действует на разных фазах развития.
Проблема эпистаза и GxE
Взаимодействие генов с окружающей средой (GxE) происходит, когда фенотипический эффект данного генетического изменения зависит от параметров окружающей среды. Сходным образом, эпистаз или взаимодействие GxG возникает, когда фенотипический эффект данного генетического изменения зависит от аллельного состояния по крайней мере одного другого локуса (Phillips, 2008; Hansen, 2013).Появляется все больше доказательств того, что взаимодействия GxG и GxE имеют фундаментальное значение для понимания эволюции и наследования сложных признаков (Gilbert and Epel, 2009; Hansen, 2013). Мы предполагаем, что оба явления могут быть интегрированы в базовую дифференциальную структуру GP, где взаимодействия GxG и GxE вводят уровень контекстной зависимости и приводят к различиям, встроенным в различия.
Разница в цветовой пигментации между темными и светлыми пляжными мышами, упомянутая ранее (рис. 3), связана не только с мутациями в Agouti , но и с кодирующей мутацией в гене MC1R , который уменьшает пигментацию (Steiner et al. , 2007; Рисунок 4Б). Эффект мутации MC1R виден только в присутствии ассоциированного со светлым цветом производного гаплотипа Agouti . Здесь считается, что локус Mc1R эпистатически взаимодействует с локусом Agouti . В этом случае мы предполагаем, что отношение GP включает не одно фенотипическое различие, а два 90 318 возможных 90 319 фенотипических различий (изменение пигментации шерсти или полное отсутствие изменений). Выбор между этими двумя фенотипическими различиями определяется генетическим фоном (здесь локус Agouti ).Дифференциальный взгляд, таким образом, остается относительно простым для взаимодействия двух локусов: зависимость фенотипа от контекста транслируется в выбор между двумя возможными фенотипическими различиями. Мы предполагаем, что отношения GP, включающие мутацию, подверженную множественным эпистатическим взаимодействиям, должны включать все возможные фенотипические различия, которые могут возникнуть в результате мутации во всех генетических фонах. Среди всех возможных фенотипических вариаций фенотипическое различие, которое будет наблюдаться, определяется другими генетическими локусами.В общем, взаимодействия GxG включают несколько сайтов, разбросанных по всему геному (Bloom et al., 2013).
РИСУНОК 4. Взаимодействие генов с окружающей средой (GxE) и GxG. (A) Мутация, кодирующая npr-1 , влияет на агрегационное поведение нематод при уровне кислорода 21%, но не при 10% (Andersen et al., 2014). (B) Мутация, кодирующая Mc1R , влияет на пигментацию тела мыши в присутствии доминантных светлых аллелей Agouti , но не в гомозиготном фоне Agouti по рецессивному темному аллелю (Steiner et al., 2007).
Примером взаимодействия GxE (см. также рис. 4A) является встречающийся в природе аллель с потерей функции brx в растениях Arabidopsis , который связан с ускоренным ростом и повышенной приспособленностью к кислым почвам, а также с сильно ослабленной корневой системой. роста по сравнению с диким типом в нормальных почвах (Gujas et al., 2012). Взаимодействия GxE обычно анализируют в форме нормы реакции , которая представляет все наблюдаемые черты одного генотипа в различных средах (Johannsen, 1911; Sarkar, 1999).В случае взаимодействий GxE мы предполагаем, что отношения GP должны включать все возможные фенотипические изменения , которые могут быть вызваны ассоциированными генетическими изменениями в различных экспериментальных условиях. Таким образом, связанное с этим фенотипическое изменение представляет собой разницу между двумя нормами реакции. Примером из учебника является изменение правила температуры и размера у C. elegans . Как и большинство других животных, нематоды C. elegans увеличиваются в размерах при низкой температуре, но лабораторный штамм дикого типа C.elegans , происходящий с Гавайских островов, не показывает изменений в размерах тела при различных температурах. Аминокислотное изменение в кальций-связывающем белке ответственно за снижение способности гавайского штамма увеличиваться в размерах при низкой температуре (Kammenga et al. , 2007). Здесь норма реакции (представляющая размер тела нематоды в диапазоне температур) различается между нематодами, и связанное с ней отношение GP охватывает разницу между этими двумя наклонами.
Диапазон фенотипических вариаций, воплощенных в отношениях GP, подверженных взаимодействиям GxG и GxE, может быть весьма огромным, особенно в случаях, когда одна и та же мутация затрагивает несколько тканей, и когда на фенотипическую изменчивость каждой ткани влияют другие геномные локусы и условия окружающей среды.Фактически, фенотипические эффекты мутации всегда зависят от других фрагментов ДНК из того же генома, так что можно считать, что любые отношения GP испытывают эпистаз. Другими словами, генетический локус, влияющий на фенотип, никогда не действует независимо от других последовательностей ДНК. Например, данный аллель опсина будет приводить к определенным свойствам цветового зрения только в том случае, если глаз сформирован и если этот глаз получает свет во время своего развития, что позволяет сформировать эффективные зрительные нейронные цепи. Для того чтобы дифференцированный взгляд был приемлемым, мы советуем не рассматривать все возможные генетические предпосылки и условия окружающей среды, а ограничивать возможности потенциальной средой и выделять аллели, которые имеют отношение к интересующей популяции (Sober, 1988).
Таким образом, при наличии эпистаза или взаимодействия GxE генетическое изменение связано не с одним фенотипическим отличием, а с несколькими возможными фенотипическими различиями, среди которых будет достигнуто одно, в зависимости от окружающей среды и генетического фона. Зависимость от контекста может быть схематически представлена как различия ГП, встроенные в другие различия генотипа и среды.
Дифференциальный взгляд на генетические и экологические эффекты на фенотипы
Как подчеркивали многие авторы (прежде всего Waddington, 1957; Oyama, 2000; Keller, 2010), гены и среда действуют на фенотип совместно, и в большинстве случаев невозможно отделить влияние одного от другого.Здесь мы показываем, что рассуждения с точки зрения различий помогают прояснить сравнение между генетическими и экологическими эффектами на фенотипы. Однако мы выделяем некоторые случаи, когда сравнение остается затруднительным.
По аналогии с отношением ГП мы можем определить отношение среда-фенотип как вариацию среды (две среды), связанное с ней фенотипическое изменение (различные фенотипические состояния) и их взаимосвязь. Например, у многих видов черепах изменение температуры во время развития яиц связано с различием полов между самцами и самками (рис. 5А), и в филогенезе черепах произошло по крайней мере шесть переходов от определения пола в зависимости от окружающей среды к генетическому (рис. 5В) (Pokorná). и Кратохвиль, 2009).В этом случае средовые и генетические эффекты можно сопоставить: половые хромосомы и температура оказывают одинаковое фенотипическое влияние на черепах. Такие наблюдения привели Уэст-Эберхард (West-Eberhard, 2003, 2005) к предложению гипотезы «гены как последователи», которая предполагает, что новые фенотипические состояния с большей вероятностью возникают в первую очередь из-за изменения окружающей среды, чем из-за генетической мутации, и что мутации происходят только позже, при изменении порога экспрессии нового признака. Уэст-Эберхард (2003, 2005) экстраполировал различия, сегрегирующие внутри популяций (различие № 2), на различия, возникшие во времени во время эволюции популяции (различие № 1).
РИСУНОК 5. Связь между средой и фенотипом в сравнении с зависимостью GP для определения пола у черепах. (A) У некоторых видов температура во время эмбрионального развития определяет пол взрослой особи. (B) У других пол определяется половыми хромосомами.
Независимая эволюция направленной лево-правой асимметрии от симметричных предков во многих линиях предоставила главный аргумент в поддержку гипотезы «ген как последователь» (Palmer, 2004).В рамках этой схемы оговаривается, что направленная асимметрия, при которой все люди односторонние, часто развилась из состояния «случайной асимметрии», когда направленность будет зависеть от факторов окружающей среды и, таким образом, различаться между генетически идентичными людьми. Например, самая сильная клешня омара будет развиваться в зависимости от использования и имеет априори равные вероятности развития с левой или с правой стороны. Мы можем видеть, как здесь работает формула «гены как последователи»: окружающая среда запускает асимметрию, и позже в ходе эволюции некоторые генетические эффекты могут изменить ее направленность в ту или иную сторону.Но в то время как асимметрия «происходит до того, как существует генетическая изменчивость, чтобы контролировать ее», дифференциальная точка зрения ясно дает понять, что генетическое влияние на направленность несопоставимо с влиянием окружающей среды, которое запускает асимметрию. Генетическое изменение приводит к переключению между окончательным 100% односторонним состоянием и начальным состоянием, при котором 50% случаев являются правосторонними и 50% левосторонними. Напротив, два альтернативных фенотипических состояния, возникающие в результате изменчивости в окружающей среде, считаются 100% правосторонним и 100% левосторонним.Этот пример показывает, что ради точности важно явно указать различия, которые учитываются в отношениях GP.
Дифференциальное представление обеспечивает теоретическую основу, которая может помочь в разработке экспериментов для исследования соответствующих переменных: можно сравнивать разные генотипы в фиксированной среде (классическая взаимосвязь GP), сравнивать реакцию фиксированного генотипа на две разные среды (фенотипическая пластичность). , или сравнить чувствительность двух разных генотипов к двум разным средам (где фенотипическая вариация становится различием в различии; см., например, Engelman et al., 2009; Томас, 2010).
Были разработаны различные количественные методы, чтобы отделить генетику от эффектов окружающей среды и количественно оценить взаимодействие GxE (Lynch and Walsh, 1998). Тем не менее, в определенных ситуациях может оказаться невозможным биологически значимым образом отделить генетические эффекты от воздействий окружающей среды, даже если рассуждать в терминах различий (Lewontin, 1974b). Популяции жука Calathus melanocephalus представлены двумя морфами — длиннокрылой и короткокрылой (Schwander, Leimar, 2011).Длиннокрылая морфа развивается только из гомозиготных особей по рецессивному аллелю, сегрегирующему в популяции, и только при хороших пищевых условиях. В этом случае генетические и экологические эффекты смешиваются (рис. 6А, В). В теоретическом случае популяции, состоящей только из короткокрылых гетерозиготных животных, выращенных в условиях голодания, и длиннокрылых, за различие крыльев между особями ответственны как гены, так и среда, и невозможно оценить соотношение факторов среды и среды. генетические эффекты, поскольку гены и окружающая среда действуют на различных уровнях сложной причинно-следственной связи между генотипами и фенотипами.
РИСУНОК 6. Перспективы взаимосвязи среда-фенотип и отношения GP для полиморфизма длины крыла у жука Calathus melanocephalus . (A) С точки зрения взаимосвязи среда-фенотип изменение пищевых условий связано с изменением размера крыла, но только в гомозиготном фоне по рецессивному аллелю ( l ) локуса размера крыла. (B) С точки зрения взаимосвязи GP, генетическое изменение в локусе размера крыла связано с изменением размера крыла, но только при хороших условиях питания.
Другой случай, который ставит под сомнение классическое различие между окружающей средой и генетикой, — это когда добавление определенных симбиотических бактерий изменяет фенотип хозяина. Мыши, которых кормили штаммом бактерий Lactobacillus , демонстрируют меньшее поведение, связанное с тревогой, по сравнению с контрольными мышами, которых кормили бульоном без бактерий (Bravo et al. , 2011). Здесь разница в поведении вызвана переключением между присутствием или отсутствием определенного кишечного симбионта. Причиной фенотипических различий является не простое изменение в последовательности ДНК, не простое изменение окружающей среды, не связанное с генетическими изменениями, а переключение между присутствием и отсутствием фактора, который можно рассматривать как фактор окружающей среды, — бактерий, — который содержит ДНК, мутации которой могут также изменить фенотип хозяина.
В заключение, рассуждения с точки зрения различий могут помочь прояснить сравнение между генетическими и экологическими эффектами на фенотипы. Тем не менее, вопросы не более чем простые. Поскольку гены и среда действуют на разных уровнях сложной причинной связи между генотипами и фенотипами, в некоторых случаях невозможно распутать обе причины.
Разъяснение терминологии «прибыль/убыток» и «разрешительный/инструктивный»
Фенотипические различия, по-видимому, подпадают под две основные категории: либо наличие/отсутствие чего-либо (например, волосы на теле или способность переваривать молоко), либо сдвиг между двумя альтернативами, которые присутствуют одновременно (например, два цвета волос). Точно так же со стороны генотипа мутация может соответствовать присутствию/отсутствию соответствующей последовательности ДНК или полиморфизму нуклеотидов. Дифференциальная точка зрения делает очевидным, что потеря фенотипа не обязательно связана с потерей генетического материала, и наоборот. Например, эволюционное приобретение темных пигментов, покрывающих всю шерсть животных, часто связывают с утратой гена Mc1R (Gompel, Prud’homme, 2009). Кроме того, как ранее отмечал один из нас (приложение Stern and Orgogozo, 2008), усиление или потеря фенотипа субъективны.Например, выпадение волос также можно рассматривать как приобретение обнаженного эпидермиса. Большинство эпидермальных клеток насекомых дифференцируются в одно из этих двух альтернативных состояний, и в обоих состояниях участвуют большие сети регуляции генов. Неясно, какое фенотипическое состояние представляет собой выигрыш или потерю по сравнению с другим. Даже с точки зрения генотипа определение потерь и приобретений может быть затруднено. Вставка мобильного элемента может сбить ген, тогда как делеция иногда может создать новый сайт связывания для активатора транскрипции.На самом деле, эволюционное усиление экспрессии desatF у D. melanogaster произошло за счет серии из трех делеций, каждая из которых создает гексамерный мотив, необходимый для экспрессии desatF (Shirangi et al., 2009).
Точно так же дифференциальная точка зрения на воздействие окружающей среды выдвигает на первый план ошибочность различия между разрешительными и инструктивными сигналами. Разрешающий сигнал связан с наличием/отсутствием фенотипа, а инструктивный сигнал — со сдвигом между двумя присутствующими альтернативами.Как утверждалось выше, эти различия на фенотипическом уровне не являются четкими.
В заключение мы предлагаем с осторожностью использовать термины «прибыль/потеря» и «поучительная/разрешительная» терминология.
Таксономически устойчивые связи ГП
Ожидается, что мутация приведет к некоторой воспроизводимой фенотипической вариации в популяции. Такая воспроизводимость фенотипического исхода необходима для обеспечения генетической эволюции и адаптации путем естественного отбора (Lewontin, 1974a; Kirschner and Gerhart, 1998).В самом деле, вновь образованный аллель, который будет генерировать еще один фенотип каждый раз, когда он попадает в другой организм, не будет подвергаться естественному отбору. Рассуждение с точки зрения изменчивости, а не рассмотрение аллелей как изолированных объектов, проясняет, что конкуренция происходит между аллелями, которые охватывают один и тот же генетический локус. Естественный отбор воздействует непосредственно на аллельную вариацию, которая устойчиво связана с данной фенотипической вариацией, что и является родством ГП. Таким образом, отношения ГП являются базовой единицей эволюционных изменений, на которые действует естественный отбор.
Главное открытие последних 20 лет состоит в том, что изменчивость в определенных генетических локусах приводит к сопоставимой фенотипической изменчивости не только у разных особей одной популяции, но и у чрезвычайно разных таксонов (Martin and Orgogozo, 2013b). Другими словами, определенные отношения GP являются таксономически надежными и присутствуют у большого количества видов. Это означает, что генетические и экологические предпосылки оставались относительно постоянными или неоднократно появлялись на протяжении эволюции, что позволяло генетическим локусам генерировать сходные фенотипические изменения в различных таксономических группах.Это важное открытие было совершенно неожиданным около 50 лет назад. Долгое время ожидалось, что сингулярность, наблюдаемая в живом мире, будет отражать сравнимую сингулярность на генетическом уровне, затрагивая несопоставимые и неконсервативные гены, специфичные для каждой линии (Mayr, 1963). Как однажды предположил Майр (1963) в 1963 году: «Многое из того, что было известно о физиологии генов, делает очевидным, что поиск гомологичных генов совершенно бесполезен, за исключением очень близких родственников […]. Поговорка «Многие дороги ведут в Рим» справедлива как в эволюции, так и в повседневных делах» (Mayr, 1963). Другими словами, ожидалось, что генетические локусы, которые делают человека мужчиной, будут отличаться от тех, которые делают собаку или рыбу. Позднее, в 80–90-х годах, некоторые исследователи предположили, что эволюция происходит за счет мутаций в консервативных генах, кодирующих белок (Romero-Herrera et al., 1978; Perutz, 1983; Stewart et al., 1987; Carroll et al. ., 2005) – но у них было мало экспериментальных данных, подтверждающих их точку зрения (Tautz and Schmid, 1998). Накопившиеся на сегодняшний день данные о мутациях, ответственных за естественную изменчивость, ясно показывают, что разнообразие живых организмов имеет общую генетическую основу как минимум в трех пунктах.Во-первых, сравнительная биология развития показала, что животные имеют общие наборы ключевых регуляторных генов с законсервированными функциями (Wilkins, 2002, 2014; Carroll et al., 2005). Во-вторых, большинство межвидовых различий у животных и растений, для которых хотя бы частично идентифицирована лежащая в основе генетическая основа (154 случая из 160), обусловлено мутациями в гомологичных генах, и очень немногие (6/160) обусловлены новыми генами. которые, тем не менее, представляют собой дубликаты существующих генов (Martin and Orgogozo, 2013b).В-третьих, было показано, что множественные случаи сходных фенотипических изменений связаны с мутациями одних и тех же гомологичных генов в независимых линиях (таблица 1), иногда на больших филогенетических расстояниях. Например, разница в пигментации между белыми и оранжевыми бенгальскими тиграми недавно была сопоставлена с единственной мутацией в гене белка-транспортера SLC45A2 (Xu et al., 2013), и этот ген также был связан с гипопигментацией глаз, кожи , волосы и перья у людей и кур (Xu et al., 2013; Рисунок 1Е). Более ярким примером является недавняя эволюция устойчивости к токсинам у трех видов, которые разошлись более 500 миллионов лет назад – моллюска, змеи и иглобрюха – посредством замены одной и той же аминокислоты в законсервированном гене (Bricelj et al., 2005). ; Geffeney et al., 2005; Venkatesh et al., 2005; Feldman et al., 2012). Такие поразительные паттерны генетического повторения в настоящее время обнаружены для более чем 100 генов животных и растений (Martin and Orgogozo, 2013b). Несмотря на существующие методологические предубеждения в пользу консервативных генов при поиске локусов количественных признаков (Rockman, 2012; Martin and Orgogozo, 2013b), уровень генетического повторения остается поразительным и предполагает, что для эволюции по крайней мере некоторых фенотипических различий требуется относительно мало генетических путей. ведут в Рим (Stern, 2013).В настоящее время не следует удивляться тому, что фрагмент ДНК, связанный со сложной окраской крыльев у одного из видов бабочек Heliconius , обеспечивает сходные крылья и коллективную защиту от одних и тех же хищников при введении в геном других бабочек (Supple et al., 2014). То, что делает собаку собакой или человека человеком, теперь частично объясняется уникальным набором таксономически устойчивых отношений ГП, которые обнаруживаются во многих ветвях линии.
Определенные отношения между средой и фенотипом также таксономически устойчивы.Например, у большинства таксонов на размер тела влияет питание; Дефицит железа может вызвать анемию, а некоторые токсичные соединения могут привести к летальному исходу. У экзотермов температура организма зависит от температуры окружающей среды. Учитывая устрашающее количество условий окружающей среды, которые можно представить, вероятно, невозможно определить, преобладают ли таксономически устойчивые отношения GP или таксономически устойчивые отношения среда-фенотип. Кроме того, вопрос о том, представляют ли таксономически устойчивые отношения GP исключительную и небольшую долю или значительную долю всех отношений GP, является предметом споров.В любом случае, существование таксономически устойчивых взаимосвязей GP теперь ясно и должно быть широко признано исследовательским сообществом.
Некоторые из самых поразительных учений современной биологии включают в себя открытие того, что живые существа имеют один и тот же генетический материал (ДНК или РНК), один и тот же генетический код (за некоторыми исключениями) и один и тот же базовый клеточный механизм. Таким образом, далеко не парадоксально, что индивидуальные различия строятся на сходствах, и тот факт, что определенные отношения ГП сохраняются на протяжении длительного времени эволюции, завершает картину.
Точная предсказательная сила, возникающая в результате существования таксономически устойчивых взаимосвязей GP, редко встречается в биологии и только начинает использовать свой полный потенциал. Сохранение связей GP на большие расстояния теперь полностью оправдывает использование подходов сравнительной генетики для решения прагматических проблем. Например, одомашнивание сельскохозяйственных культур приняло форму аналогичного давления отбора у многих видов, и теперь у нас есть экспериментальные доказательства того, что этот процесс неоднократно включал мутации в одном и том же наборе консервативных генов (Патерсон и др., 1995; Мартин и Оргогозо, 2013b). Это наблюдение открывает интересные возможности для применения, поскольку мы можем использовать эту новую совокупность генетических знаний, чтобы помочь одомашниванию будущих культур или использовать стратегии с помощью маркеров для производства и поддержания биоразнообразия сельскохозяйственных культур (Lenser and Theißen, 2013). Предсказуемость GP уже используется для идентификации штаммов, которые развили устойчивость к различным стратегиям борьбы с вредителями, причем крайние случаи нацелены на толерантность к противомалярийным препаратам у паразитов Plasmodium (Manske et al., 2012), устойчивость бактерий и дрожжей к антибиотикам (Fischbach, 2009; MacCallum et al., 2010) или, что еще более драматично, антропогенная эволюция устойчивости к инсектицидам у различных когорт насекомых, независимо от их статуса вредителя (Ffrench-Constant и др., 2004; Мартин и Оргогозо, 2013b).
Кроме того, повторяемость генетической основы фенотипической изменчивости предполагает, что клинические исследования, вероятно, также получат пользу от генетических исследований большого числа модельных видов (Robinson and Webber, 2014).Например, естественная изменчивость толерантности к химиотерапевтическому препарату метотрексату была картирована у плодовых мушек Drosophila с генами, человеческие ортологи которых также связаны с реакцией пациентов на этот препарат (Кислухин и др. , 2013), что расширило использование модельных организмов в качестве моделей болезней.
На пути к генной классификации фенотипов
Одним из оригинальных аспектов описания связи GP с точки зрения индивидуальных отношений GP является то, что это позволяет классифицировать фенотипы в соответствии с лежащей в их основе генетической основой.На первом уровне отношения GP, затрагивающие разные регионы одного и того же гена и дающие сопоставимые фенотипические результаты, могут быть сгруппированы вместе. Простые случаи взаимодействия GxG были обнаружены между тесно сцепленными мутациями, как правило, внутри кодирующей последовательности или внутри регуляторного элемента цис-, когда они вызывают неаддитивный эффект на фенотип. Например, было обнаружено, что определенная мутация в энхансере вызывает различные сдвиги в паттерне экспрессии нижестоящего кодирующего гена в зависимости от соседней последовательности ДНК (Frankel et al., 2011; Роджерс и др., 2013). Сходным образом было обнаружено, что аминокислотные мутации в гене гемоглобина увеличивают или уменьшают сродство к кислороду в зависимости от аллельного состояния других сайтов (Natarajan et al. , 2013). В таких случаях интуитивно понятно сгруппировать такие генетически связанные сайты вместе, поскольку все они влияют на один и тот же тип фенотипического признака.
Отсутствие пигментов меланина у животных было связано с мутациями в нескольких генах, включая OCA2 , kit-лиганд или Mc1R (рассмотрено в Gompel and Prud’homme, 2009; Liu et al., 2013). В то время как отсутствие меланина традиционно рассматривается как одно из состояний признака, альбинизм, независимо от лежащей в основе генетической основы, мы предлагаем здесь различать OCA2 -ассоциированный альбинизм от Mc1R -ассоциированного альбинизма или от альбинизма, связанного с любым другим геном. Одним из преимуществ разложения изменчивости живого мира на эти множественные элементарные отношения ЗП является то, что эти элементы затем могут быть сгруппированы в последовательно увеличивающиеся группы.Элементарные фенотипические изменения, затрагивающие разные гены, которые являются частью одного и того же генетического пути, также могут быть сгруппированы вместе как сопутствующие компоненты одного и того же механизма модуляции фенотипа. Это явно относится к сигнальным молекулам TGF-β BMP15, GDF9 и рецептору TGF-β BMPR1B, которые неоднократно связывались с изменениями функции яичников у людей и пород домашних овец (обзор в Luong et al., 2011).
Другим важным следствием перспективы родства ГП является то, что очевидно различные фенотипические изменения, вызванные сходными генетическими локусами в различных организмах, могут быть исследованы далее, чтобы выявить то, что может быть общим основным фенотипическим изменением (Deans et al., 2015). Например, личинки мух и черви-нематоды имеют разное поведение при поиске пищи, но было показано, что мутации в одном и том же ортологическом гене ( вместо / egl-4 ) изменяют интенсивность поведения при поиске пищи у обоих организмов (Osborne et al. , 1997; Мери и др., 2007; Хонг и др., 2008). Таким образом, вполне вероятно, что основное поведенческое изменение, которое лежит в основе кажущихся отличительными изменений поиска пищи мухами и нематодами, представляет собой консервативное отношение GP между нематодами и мухами. Этот несколько пограничный пример иллюстрирует проблему включения широко распространенного сравнительного мышления в наше глобальное понимание биологии. Имеет ли отношение мутация в мышиной модели к заболеванию человека? Можем ли мы считать, что фенотип мыши подобен состоянию человека, если его генетическая основа иная? Мы и другие прогнозируем, что поиск ортологичных фенотипов, или «фенологов» (McGary et al., 2010), станет главной задачей современной генетики и потребует плодотворного союза между прикладной и эволюционной биологией.
Заключение
В этой статье мы возвращаем дифференциальную концепцию гена (Schwartz, 2000) в нашу структуру для понимания карты GP. Дифференциальный взгляд на отношения GP помогает прояснить генетические и экологические эффекты на фенотипы и их связь. Это также открывает новые возможности для размышлений, в частности, в отношении декомпозиции наблюдаемых признаков внутри организма и представления карт GP. Кроме того, существование таксономически надежных взаимосвязей GP поощряет беззастенчивое использование сравнительной генетики для предсказания генетической основы фенотипической изменчивости в различных группах организмов, и эта предсказательная сила имеет важный потенциал для трансляционных исследований в области агрономии и клинических исследований.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы глубоко признательны Мари-Анн Феликс за содержательные обсуждения и Томасу Прадеу за то, что он привлек наше внимание к нескольким важным документам. Мы также благодарим Giuseppe Baldacci, Marie-Anne Félix, Pierre-Henri Gouyon, Alexandre Peluffo, Mark Siegal, David Stern и рецензентов за их проницательные комментарии к рукописи.Исследование, предшествовавшее написанию этой статьи, финансировалось Европейским исследовательским советом в рамках Седьмой рамочной программы Европейского сообщества (соглашение о гранте FP7/2007-2013 № 337579) и Фондом Джона Темплтона (грант № 43903).
Сокращения
GP, генотип-фенотип.
Ссылки
Андерсен, Э. К., Блум, Дж. С., Герке, Дж. П., и Кругляк, Л. (2014). Вариант нейропептидного рецептора npr-1 является основной детерминантой роста и физиологии Caenorhabditis elegans . Генетика PLoS. 10:e1004156. doi: 10.1371/journal.pgen.1004156
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Блум, Дж. С., Эренрайх, И. М., Лоо, В. Т., Лайт, Т.-Л. В. и Кругляк Л. (2013). Поиск источников отсутствующей наследственности при скрещивании дрожжей. Природа 494, 234–237. doi: 10.1038/nature11867
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бойкот, А. Э., Дайвер, К., Гарстанг, С. Л., и Тернер, Ф.М. (1931). Наследование левосторонней формы у limnæa peregra (Mollusca, Pulmonata). Филос. Транс. Р. Соц. Лонд. сер. В 219, 51–131. doi: 10.1098/rstb.1931.0002
Полнотекстовая перекрестная ссылка | Академия Google
Браво, Дж. А., Форсайт, П., Чу, М. В., Эскаравэдж, Э., Савиньяк, Х. М., Динан, Т. Г., и соавт. (2011). Прием внутрь штамма Lactobacillus регулирует эмоциональное поведение и экспрессию центрального рецептора ГАМК у мышей через блуждающий нерв. Проц.Натл. акад. науч. США 108, 16050–16055. doi: 10.1073/pnas.1102999108
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Bricelj, V.M., Connell, L., Konoki, K., Macquarrie, S.P., Scheuer, T., Catterall, W.A., et al. (2005). Мутация натриевого канала, приводящая к устойчивости моллюсков к сакситоксину, увеличивает риск ПНП. Природа 434, 763–767. doi: 10.1038/nature03415
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кэрролл, С.Б., Гренье Дж. и Уэзерби С. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных . Молден, Массачусетс: John Wiley & Sons.
Академия Google
Кассирер, Э. (1910). Вещество и функция , 2010 Edn. Париж: Набу Пресс.
Академия Google
Коэн, Э. (2012). От клеток к цивилизациям: принципы изменения, формирующие жизнь . Принстон, Нью-Джерси: Издательство Принстонского университета. дои: 10.1515/9781400841653
Полнотекстовая перекрестная ссылка | Академия Google
Колозимо, П.Ф., Хосеманн, К.Е., Балабхадра, С., Вильяреал, Г. мл., Диксон, М., Гримвуд, Дж., и соавт. (2005). Широко распространенная параллельная эволюция у колюшек путем повторной фиксации аллелей эктодисплазина. Наука 307, 1928–1933. doi: 10.1126/science.1107239
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Докинз, Р. (1982). Расширенный фенотип: длинная досягаемость гена . Оксфорд: Издательство Оксфордского университета.
Академия Google
Динс, А.R., Lewis, S.E., Huala, E., Anzaldo, S.S., Ashburner, M., Balhoff, J.P., et al. (2015). Находим свой путь через фенотипы. PLoS Биол. 13:e1002033. doi: 10.1371/journal.pbio.1002033
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Данбар, Х.Е., Уилсон, А.К.С., Фергюсон, Н.Р., и Моран, Н.А. (2007). Термическая устойчивость тли определяется точечной мутацией в бактериальных симбионтах. PLoS Биол. 5:e96. doi: 10.1371/journal.pbio.0050096
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Энгельман, К.D., Baurley, J.W., Chiu, Y.-F., Joubert, B.R., Lewinger, J.P., Maenner, M.J., et al. (2009). Обнаружение взаимодействий генов и окружающей среды в данных полногеномной ассоциации. Жен. Эпидемиол. 33(Прил. 1), S68–S73. doi: 10.1002/gepi.20475
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Feldman, C.R., Brodie, E.D. Jr., Brodie, E.D. III, and Pfrender, ME (2012). Ограничение формирует конвергенцию в устойчивых к тетродотоксину натриевых каналах змей. Проц.Натл. акад. науч. США 109, 4556–4561. doi: 10.1073/pnas.1113468109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Франкель, Н., Эрезилмаз, Д. Ф., МакГрегор, А. П., Ван, С., Пайр, Ф., и Стерн, Д. Л. (2011). Морфологическая эволюция, вызванная множеством малозаметных замен в регуляторной ДНК. Природа 474, 598–603. doi: 10.1038/nature10200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Геффени, С.Л., Фуджимото, Э., Броди, Э.Д. III, Броди, Э.Д. младший, и Рубен, П.С. (2005). Эволюционная диверсификация устойчивых к ТТХ натриевых каналов во взаимодействии хищник-жертва. Природа 434, 759–763. doi: 10.1038/nature03444
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gerstein, M.B., Bruce, C., Rozowsky, J.S., Zheng, D., Du, J., Korbel, J.O., et al. (2007). Что такое ген после кодирования? История и обновленное определение. Рез. генома. 17, 669–681.doi: 10.1101/gr.6339607
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гилберт, С. Ф. (2000). «Классические гены и гены развития: различное использование генов в эволюционном синтезе», в The Concept of the Gene in Development and Evolution , eds P. Beurton, R. Falk, and H.-J. Райнбергер (Кембридж, Нью-Йорк: издательство Кембриджского университета).
Гилберт, С. Ф., и Эпель, Д. (2009). Экологическая биология развития: интеграция эпигенетики, медицины и эволюции .Сандерленд, Массачусетс: Sinauer Associates.
Академия Google
Гьювсланд, А.Б., Вик, Дж.О., Берд, Д.А., Хантер, П.Дж., и Омхолт, С.В. (2013). Преодоление разрыва между генотипом и фенотипом: что для этого нужно? J. Physiol. 591, 2055–2066. doi: 10.1113/jphysiol.2012.248864
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гриффитс, П., и Стоц, К. (2013). Генетика и философия: введение . Кембридж: Издательство Кембриджского университета.дои: 10.1017/CBO9780511744082
Полнотекстовая перекрестная ссылка | Академия Google
Гуджас, Б., Алонсо-Бланко, К., и Хардтке, К.С. (2012). Природные аллели потери функции Arabidopsis brx обеспечивают адаптацию корней к кислой почве. Курс. биол. 22, 1962–1968. doi: 10.1016/j.cub.2012.08.026
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холлочер, Х. , Хэтчер, Дж. Л., и Дайресон, Э. Г. (2000). Генетический и онтогенетический анализ различий в пигментации брюшка у разных видов подгруппы Drosophila dunni . Эволюция. Междунар. Дж. Орг. Эвол. 54, 2057–2071. doi: 10.1111/j.0014-3820.2000.tb01249.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хонг, Р. Л., Витте, Х., и Соммер, Р. Дж. (2008). Естественная вариация в привлечении феромонов насекомых Pristionchus pacificus включает протеинкиназу EGL-4. Проц. Натл. акад. науч. США 105, 7779–7784. doi: 10.1073/pnas.0708406105
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джонс, Ф.C., Grabherr, M.G., Chan, Y.F., Russell, P., Mauceli, E., Johnson, J., et al. (2012). Геномные основы адаптивной эволюции трехиглой колюшки. Природа 484, 55–61. doi: 10.1038/nature10944
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Камберов Ю.Г. , Ван С., Тан Дж., Жербо П., Варк А., Тан Л. и соавт. (2013). Моделирование недавней эволюции человека у мышей путем экспрессии выбранного варианта EDAR. Сотовый 152, 691–702. дои: 10.1016/j.cell.2013.01.016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Камменга, Дж. Э., Доросук, А., Риксен, Дж. А. Г., Хазендонк, Э., Спиридон, Л., Петреску, А.-Дж., и соавт. (2007). Caenorhabditis elegans дикого типа не поддается правилу «температура-размер» из-за полиморфизма одного нуклеотида в tra-3. PLoS Жене 3:e34. doi: 10.1371/journal.pgen.0030034
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Келлер, Э.Ф. (2010). Мираж пространства между природой и воспитанием . Дарем, Северная Каролина: Издательство Университета Дьюка. дои: 10.1215/9780822392811
Полнотекстовая перекрестная ссылка | Академия Google
Киршнер М. и Герхарт Дж. (1998). Эволюционируемость. Проц. Натл. акад. науч. США 95, 8420–8427. doi: 10.1073/pnas.95.15.8420
Полнотекстовая перекрестная ссылка | Академия Google
Кислухин Г., Кинг Э. Г., Уолтерс К. Н., Макдональд С. Дж. и Лонг А. Д. (2013). Генетическая архитектура токсичности метотрексата сходна у Drosophila melanogaster и человека.3, 1301–1310. doi: 10.1534/g3.113.006619
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ландри, Ч.Р., и Рифкин, С.А. (2012). Генотип-фенотипические карты системной биологии и количественной генетики: различные и взаимодополняющие. Доп. Эксп. Мед. биол. 751, 371–398. дои: 10.1007/978-1-4614-3567-9_17
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ланглад, Н. Б., Фэн, X., Дрансфилд, Т., Копси, Л., Ханна, А. И., Thebaud, C., et al. (2005). Эволюция через генетически контролируемое аллометрическое пространство. Проц. Натл. акад. науч. США 102, 10221–10226. doi: 10.1073/pnas. 0504210102
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Левонтин Р. (1974а). Генетическая основа эволюционных изменений (Колумбийская биологическая серия) . Колумбия: Издательство Колумбийского университета.
Академия Google
Левонтин, Р. К. (1974b). Дисперсионный анализ и анализ причин. утра. Дж. Хам. Жене. 26, 400–411.
Академия Google
Linnen, C.R., Poh, Y.-P., Peterson, B.K., Barrett, R.D.H., Larson, J.G., Jensen, J.D., et al. (2013). Адаптивная эволюция нескольких признаков посредством множественных мутаций в одном гене. Наука 339, 1312–1316. doi: 10.1126/science.1233213
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Luong, H.T.T., Chaplin, J., McRae, A.F., Medland, S.E., Willemsen, G., Nyholt, D.R., et al. (2011). Вариации BMPR1B, TGFRB1 и BMPR2 и контроль дизиготного двойникования. Двойной рез. Гум. Жене. Выключенный. Дж. Междунар. соц. Твин Стад. 14, 408–416. doi: 10.1375/twin.14.5.408
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Линч М. и Уолш Б. (1998). Генетика и анализ количественных признаков . Сандерленд, Массачусетс: Sinauer Associates.
Академия Google
МакКаллум, Д. М., Кост, А., Ишер, Ф., Якобсен, М. Д., Оддс, Ф. К., и Санглард, Д. (2010). Генетическое исследование механизмов устойчивости к азолам у Candida albicans и их проверка на модели диссеминированной инфекции на мышах. Антимикроб. Агенты Чемотер. 54, 1476–1483. doi: 10.1128/AAC.01645–1649
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Манске М., Миотто О., Кампино С., Оберн С., Альмагро-Гарсия Дж., Маслен Г. и др. (2012). Анализ разнообразия Plasmodium falciparum при естественных инфекциях методом глубокого секвенирования. Природа 487, 375–379. doi: 10.1038/nature11174
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мартин А. и Оргогозо В. (2013b). Локусы повторной эволюции: каталог генетических очагов фенотипической изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 67, 1235–1250. doi: 10.1111/evo.12081
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
МакГэри, К.Л., Парк, Т.Дж., Вудс, Дж.О., Ча, Х.Дж., Уоллингфорд, Дж.Б. и Маркотт, Э. М. (2010). Систематическое открытие неочевидных моделей болезней человека с помощью ортологических фенотипов. Проц. Натл. акад. науч. США 107, 6544–6549. doi: 10.1073/pnas.0
0107
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мери, Ф., Белай, А.Т., Со, А.К.-С., Соколовский, М.Б., и Кавецкий, Т.Дж. (2007). Природный полиморфизм, влияющий на обучение и память у дрозофилы . Проц. Натл. акад. науч. США 104, 13051–13055.doi: 10.1073/pnas.0702923104
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Натараджан С., Иногути Н. , Вебер Р. Э., Фаго А., Морияма Х. и Сторц Дж. Ф. (2013). Эпистаз среди адаптивных мутаций гемоглобина оленьих мышей. Наука 340, 1324–1327. doi: 10.1126/science.1236862
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ноубл, Д. (2006). Музыка жизни: биология за пределами генома . Оксфорд: Издательство Оксфордского университета.
Академия Google
Осборн, К. А., Робишон, А., Берджесс, Э., Батланд, С., Шоу, Р. А., Култхард, А., и соавт. (1997). Естественный полиморфизм поведения, обусловленный цГМФ-зависимой протеинкиназой Drosophila . Наука 277, 834–836. doi: 10.1126/наука.277.5327.834
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ояма, С. (2000). Онтогенез информации: системы развития и эволюция , 2-е изд.Кембридж, Нью-Йорк: Издательство Университета Дьюка. дои: 10.1215/9780822380665
Полнотекстовая перекрестная ссылка | Академия Google
Патерсон, А. Х., Лин, Ю. Р., Ли, З., Шерц, К. Ф., Добли, Дж. Ф., Пинсон, С. Р., и соавт. (1995). Конвергентное одомашнивание злаковых культур путем независимых мутаций в соответствующих генетических локусах. Наука 269, 1714–1718. doi: 10.1126/наука.269.5231.1714
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Покорна, М., и Кратохвил, Л. (2009). Филогения механизмов определения пола у чешуйчатых рептилий: половые хромосомы — эволюционная ловушка? Зоол. Дж. Линн. соц. 156, 168–183. doi: 10.1111/j.1096-3642.2008.00481.x
Полнотекстовая перекрестная ссылка | Академия Google
Рокман, М. В. (2012). Программа QTN и аллели, имеющие значение для эволюции: все золото не блестит. Эволюция. Междунар. Дж. Оргн. Эвол. 66, 1–17. doi: 10.1111/j.1558-5646.2011.01486.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Роджерс, В. А., Саломоне, Дж. Р., Тейси, Д. Дж., Камино, Э. М., Дэвис, К. А., Ребейс, М., и соавт. (2013). Рекуррентная модификация консервативного цис-регуляторного элемента лежит в основе разнообразия пигментации плодовых мушек. PLoS Genet 9:e1003740. doi: 10.1371/journal.pgen.1003740
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ромеро-Эррера, А.E., Lehmann, H., Joysey, K.A., и Friday, A.E. (1978). Об эволюции миоглобина. Филос. Транс. Р. Соц. Лонд. Б. биол. науч. 283, 61–163. doi: 10.1098/rstb.1978.0018
Полнотекстовая перекрестная ссылка | Академия Google
Саркар, С. (1999). От нормы реакции к норме адаптации: норма реакции, 1909–1960 гг. Биол. Филос. 14, 235–252. дои: 10.1023/A:10066648
Полнотекстовая перекрестная ссылка | Академия Google
Шварц, С.(2000). «Дифференциальная концепция гена: прошлое и настоящее», в The Concept of the Gene in Development and Evolution, eds PJ Beurton, R. Falk and HJ Rheinberger (Cambridge, NY: Cambridge University Press), 26–39. . дои: 10.1017/CBO9780511527296.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ширанги, Т. Р., Дюфур, Х. Д., Уильямс, Т. М., и Кэрролл, С. Б. (2009). Быстрая эволюция экспрессии фермента, продуцирующего половые феромоны, у Drosophila . PLoS Биол. 7:e1000168. doi: 10.1371/journal.pbio.1000168
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Симондон, Г. (1968). L’individuation à la Lumière des Notions de Forme et d’Information , 2005th Edn. Гренобль: Миллон.
Академия Google
Steiner, C.C., Weber, JN, and Hoekstra, H.E. (2007). Адаптивная вариация у пляжных мышей, вызванная двумя взаимодействующими генами пигментации. PLoS Биол. 5:e219.doi: 10.1371/journal.pbio.0050219
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стерн, Д.Л. (2000). Эволюционная биология развития и проблема изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 54, 1079–1091. doi: 10.1111/j.0014-3820.2000.tb00544.x
Полнотекстовая перекрестная ссылка | Академия Google
Стоц, К. (2012). Убийство на экспрессе развития: кто убил природу/воспитание? Биол . Филос. 27, 919–929.doi: 10.1007/s10539-012-9343-1
Полнотекстовая перекрестная ссылка | Академия Google
Стертевант, А. Х. (1932). Использование мозаики в изучении влияния генов на развитие. Проц. Шестой межд. конгр. Жене. Итака, штат Нью-Йорк, 1, 304–307.
Саппл, М., Папа, Р., Контрман, Б., и Макмиллан, В. О. (2014). Геномика адаптивного излучения: понимание континуума видообразования геликониуса. Доп. Эксп. Мед. Биол . 781, 249–271. дои: 10.1007/978-94-007-7347-9_13
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Таутц, Д.и Шмид, К. Дж. (1998). От генов к индивидуумам: гены развития и формирование фенотипа. Филос. Транс. Р. Соц. Лонд. Б. биол. науч. 353, 231–240. doi: 10.1098/rstb.1998.0205
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Венкатеш, Б., Лу, С.К., Дандона, Н., См., С.Л., Бреннер, С., и Сунг, Т.В. (2005). Генетическая основа устойчивости к тетродотоксину у иглобрюхов. Курс. биол. 15, 2069–2072. doi: 10.1016/j.cub.2005.10.068
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вагнер, Г. П. (1996). Гомологи, природные виды и эволюция модульности. утра. Зоол. 36, 36–43. doi: 10.1093/icb/36.1.36
Полнотекстовая перекрестная ссылка | Академия Google
Вагнер, Г. П. (2000). Концепция характера в эволюционной биологии .Сан-Диего, Калифорния: Academic Press.
Академия Google
Вагнер, Г. П. (2014). Гомология, гены и эволюционные инновации . Принстон, Нью-Джерси: Издательство Принстонского университета. дои: 10.1515/9781400851461
Полнотекстовая перекрестная ссылка | Академия Google
Wang, J., Wurm, Y., Nipitwattanaphon, M., Riba-Grognuz, O., Huang, Y.-C., Shoemaker, D., et al. (2013). Y-подобная социальная хромосома вызывает альтернативную организацию колонии у огненных муравьев. Природа 493, 664–668.doi: 10.1038/nature11832
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уотерс, CK (2007). Причины, которые имеют значение. Дж. Филос. 104, 551–579.
Академия Google
Велтер, Д., Макартур, Дж., Моралес, Дж., Бердетт, Т., Холл, П., Джанкинс, Х., и др. (2014). Каталог NHGRI GWAS, курируемый ресурс ассоциаций SNP-признаков. Рез. нуклеиновых кислот. 42, Д1001–Д1006. doi: 10.1093/nar/gkt1229
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вест-Эберхард, М.Дж. (2003). Пластичность развития и эволюция . Оксфорд: Издательство Оксфордского университета.
Академия Google
Уилкинс, А. (2014). «Набор генетических инструментов»: история жизни важной метафоры», в Advances in Evolutionary Developmental Biology , изд. Дж. Тодд Стрилман (Хобокен, Нью-Джерси: John Wiley & Sons), 1–14.
Академия Google
Уилкинс, А.С. (2002). Эволюция путей развития . Сандерленд, Массачусетс: Sinauer Associates.
Академия Google
Xu X., Dong G.-X., Hu X.-S., Miao L., Zhang X.-L., Zhang D.-L., et al. (2013). Генетическая основа белых тигров. Курс. биол. 23, 1031–1035. doi: 10.1016/j.cub.2013.04.054
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
границ | Изменение цвета, фенотипическая пластичность и камуфляж
Введение
Одна из главных проблем, с которыми сталкиваются животные, — это как не быть съеденными хищниками.Соответственно, многие виды развили маскировку, чтобы избежать обнаружения или распознавания (Stevens and Merilaita, 2009). Однако значительная проблема заключается в том, как люди могут сопоставлять диапазон визуальных фонов, на которых их часто видят. Большинство типов камуфляжа связано с взаимодействием между окружающей средой и фенотипом животного, при этом внешний вид должен в большей или меньшей степени напоминать фон, на котором наблюдается животное. Большинство мест обитания не однородны, а демонстрируют значительное разнообразие цвета, яркости и рисунка.Одно из решений этой проблемы включает в себя отбор, благоприятствующий эволюции генетических полиморфизмов внешнего вида, когда особи попадают в одну из множества дискретных морфов. Такие полиморфизмы и их генетическая основа хорошо изучены у мышей и рептилий (Nachman et al., 2003; Rosenblum et al., 2004; Rosenblum, 2006).
КЛЮЧЕВОЕ ПОНЯТИЕ 1. полиморфизм Многие виды ракообразных, меняющих окраску, также высоко полиморфны, и это может позволить провести ценные испытания механизмов и функций камуфляжа, а также решить вопросы, касающиеся внутривидового разнообразия.
Проблема генетического полиморфизма в том, что ему не хватает гибкости; человек по-прежнему ограничен внешним видом, который обеспечил бы эффективную маскировку на ограниченном диапазоне фонов или в одном типе среды обитания. Камуфляж нельзя модифицировать, чтобы он справился с изменениями окружающей среды со временем или с высокой неоднородностью внешнего вида среди пятен в пространстве. Многие из этих проблем потенциально могут быть решены за счет способности отдельных животных менять внешний вид с течением времени. Это широко задокументировано для многочисленных видов и таксонов, от крабов и гусениц до млекопитающих и рептилий (Stuart-Fox and Moussalli, 2009; Umbers et al., 2014). Изменение цвета используется не только для маскировки. Это обычно происходит у видов для сигнальных функций, особенно выбора партнера, начиная от изменения цвета и яркости у хамелеонов (Ligon and McGraw, 2013) до сигналов поляризации у креветок-богомолов (Cronin et al., 2009). Другая важная функция, вероятно, заключается в терморегуляции. Что касается камуфляжа, то большинство исследований было сосредоточено на относительно небольшом количестве видов, в частности на тех, которые способны к чрезвычайно быстрым изменениям (секунды), особенно на головоногих (Hanlon et al., 2009; Цзяо и др., 2011 г.; Зилински и Йонсен, 2011). Хотя эти исследования показали многое о том, как изменение цвета можно использовать для соответствия различным средам, и связанную с этим визуальную обратную связь, изменение цвета с такой скоростью, вероятно, будет нетипичным по своей природе, особенно за пределами головоногих моллюсков и рыб. Вместо этого большинство животных, вероятно, меняются в течение нескольких часов, дней, недель и месяцев (Umbers et al., 2014). Это может включать модификации, происходящие в результате физиологического изменения цвета, которые часто связаны с сокращением и диспергированием окрашенного пигмента в хроматофорных клетках, долгосрочные изменения, связанные с морфологией клеточного распределения и синтеза пигмента, а также в процессе развития (Stuart-Fox and Moussalli, 2009; Амберс и др. , 2014). Большинство исследований в основном сосредоточены на физиологических процессах, не всегда рассматривают функциональные и экологические проблемы, и остается множество вопросов о механизмах и адаптивном значении изменения цвета.
КЛЮЧЕВАЯ КОНЦЕПЦИЯ 2. Изменение цвета Изменение цвета может происходить в различных временных масштабах, от секунд и минут до часов и дней. Большинство работ было сосредоточено на быстром изменении цвета, но сравнительно медленное изменение, вероятно, является обычным явлением в природе.
КЛЮЧЕВАЯ КОНЦЕПЦИЯ 3.Затраты и адаптивная ценность изменения цвета Существует много нерешенных вопросов в области изменения цвета и маскировки, в том числе о том, как именно работает изменение цвета, каковы метаболические затраты и обеспечиваемое адаптивное преимущество.
Использование крабов для изучения изменения цвета и маскировки
Возможно, наиболее широко используемой группой животных, помимо головоногих, для изучения изменения цвета и его механизмов были крабы. Прошлые работы были сосредоточены, в частности, на крабах-скрипачах, Uca .Некоторые виды имеют циркадный дневной и ночной ритм изменения окраски (преимущественно яркости), при котором особи часто становятся светлее ночью и темнее днем (Аткинс, 1926; Абрамовиц, 1937; Браун и Уэбб, 1948). Это может быть связано с изменением дисперсии черного и белого пигментов в хроматофорных клетках (Abramowitz, 1937; Brown, Webb, 1948; Fingerman, 1955, 1956; Fingerman, Yamamoto, 1967; Rao et al., 1967) и наиболее вероятно. выполнять функцию защиты крабов от интенсивного ультрафиолетового излучения (Darnell, 2012).Некоторые крабы-скрипачи также демонстрируют более высокие уровни дисперсии черного и белого пигмента на темном и светлом фоне соответственно (Brown and Sandeen, 1948; Rao et al., 1967), предположительно для маскировки. В целом, несмотря на значительную работу по изменению цвета крабов Uca , большинство из них не касалось маскировки и не фокусировалось на эколого-эволюционном значении изменения цвета. Кроме того, большая часть работы, либо в течение дневных и лунных циклов, либо на разных фонах, включала оценку изменения цвета с использованием индекса дисперсии пигмента, который показывает, насколько рассредоточенные типы пигмента кажутся человеческому глазу под микроскопом.В сравнительно небольшом количестве исследований объективно оценивали изменение окраски крабов (но см. Hemmi et al., 2006; Detto et al., 2008; Darnell, 2012) и редко их маскировку (Stevens et al., 2013, 2014a; Russell and Dierssen, 2015). ).
Недавнее исследование изменения окраски у молодых рогатых крабов-призраков ( Ocypode ceratophthalmus ) оценивало изменения цвета и яркости отдельных особей с использованием мер отражения и показателей цвета, полученных в результате анализа цифровых изображений (Stevens et al., 2013).Это показало, что крабы-призраки также имеют циркадные изменения внешнего вида, но в отличие от Uca , отдельные крабы-призраки становятся темнее и серее ночью и светлее и желтее днем (рис. 1). Эти изменения, по-видимому, позволяют им лучше соответствовать цвету и яркости песчаного пляжа в течение дня и, возможно, сливаться с тенями ночью. Кроме того, как и Uca , крабы становятся светлее на белом фоне и темнее на черном фоне, что потенциально еще больше улучшает их камуфляж.Изменения яркости не происходит, когда крабов просто помещают в темноту. Эти эксперименты показывают, как изменение цвета потенциально можно использовать для модуляции маскировки в течение нескольких часов, хотя в исследовании не моделировался внешний вид или маскировка крабов-призраков для зрения хищника. В другой работе изучалась окраска и камуфляж крабов, обнаруженных на матах Sargassum , которые, по-видимому, также демонстрируют заметные изменения внешнего вида в зависимости от субстрата (Russell and Dierssen, 2015). В этой работе моделировался внешний вид и маскировка крабов для рыб и птичьего зрения, при этом камуфляж крабов лучше противостоял дихроматической системе рыб.
Рис. 1. Левое изображение показывает изменение яркости двух береговых крабов ( Carcinus maenas ) при помещении на черный (слева) и белый (справа) фон на 2 часа . Из Stevens et al. (2014а). На правом изображении показан рогатый краб-призрак днем со светло-желтой окраской (вверху) и ночью с темно-серой окраской (внизу).
Недавно мы провели эксперименты, чтобы определить, может ли обычный береговой краб ( Carcinus maenas ) менять цвет в течение 2 часов, чтобы лучше соответствовать своему фону (Stevens et al., 2014а). Предыдущая работа Пауэлла (1962б) показала, что в течение 90-минутного периода происходили изменения в распределении черного и белого пигмента в хроматофорных клетках, когда крабов помещали на черный или белый фон. Кроме того, береговые крабы также демонстрируют изменение циркадного ритма, становясь темнее днем и светлее ночью (Powell, 1962a). Однако в этой работе не анализировалось фактическое изменение окраски как такового или индивидуальное совпадение с фоном. В наших экспериментах мы проверили молодь крабов на изменение яркости на черном или белом фоне и на изменение цвета на красном или зеленом фоне, а также смоделировали степень изменения цвета и яркости (воспринимаемой легкости) особей в зрительной системе человека. птичий хищник.Мы обнаружили, что крабы могут менять свою яркость, становясь темнее на черном фоне и светлее на белом фоне (рис. 1). Хотя существовали значительные индивидуальные различия, и изменения часто не были значительными, у некоторых людей они были достаточно значительными, чтобы привести к лучшему совпадению фона с моделью визуального различения. Будет ли это равносильно снижению риска хищничества, требует проверки с помощью экспериментов по хищничеству / обнаружению. Напротив, мы не обнаружили изменений в цвете на зеленом и красном фоне, по крайней мере, в той степени, в которой это могло бы повлиять на камуфляж.
Береговые крабы, как и ряд подобных видов, представляют собой превосходную систему для изучения изменения цвета и пластичности, а также их роли в сокрытии. Во-первых, появляется все больше свидетельств того, что у молодых особей могут происходить некоторые изменения яркости, которые могут улучшить маскировку (Powell, 1962b; Stevens et al., 2014a). Кроме того, они, весьма вероятно, претерпевают существенные изменения внешнего вида за счет фенотипической пластичности во время линьки (Todd et al., 2006; Stevens et al., 2014b; Jensen and Egnotovich, 2015; рис. 2).Действительно, паттерны передачи сигналов у крабов-скрипачей могут претерпевать существенные изменения между линьками (Detto et al., 2008). Во-вторых, береговые крабы — распространенный и широко распространенный вид, встречающийся во многих местах обитания, потенциально требующий разного внешнего вида для обеспечения маскировки в каждом из них. В соответствии с этим крабы из каменистых, илистых и мидиевых местообитаний демонстрируют различия как в цвете, так и в рисунке (Todd et al., 2006, 2012; Stevens et al., 2014b; рисунок 3). Тем не менее, стоит отметить, что, несмотря на ряд исследований, изучающих ассоциации фенотип-окружающая среда , которые, вероятно, способствуют камуфляжу (для целого ряда таксонов животных), только одно исследование песчаных блох непосредственно тестировало камуфляж (напр. г., фенотип-среда соответствует ; Стивенс и др., 2015). В-третьих, береговые крабы сильно различаются по внешнему виду даже в одном и том же месте, по-видимому, с широким спектром тактик маскировки от совпадения с фоном до разрушительной окраски (Hogarth, 1978; Stevens et al., 2014b; рис. 4). Уровень разнообразия связан с типом среды обитания / местом, где они живут. Это открытие может позволить проверить, насколько высокое внутривидовое разнообразие создается и поддерживается. В-четвертых, их биология и крепкий характер делают их идеальным видом для проведения экспериментов и определения механизмов изменения цвета.Наконец, за пределами своего естественного европейского ареала они высоко инвазивны по всему миру (один из «100 лучших» инвазивных видов в мире), а это означает, что они представляют собой ценную систему для изучения биологии инвазии (Darling et al., 2008; McGaw et al. ., 2011).
КЛЮЧЕВОЕ ПОНЯТИЕ 4. Линька и пластичность Многие крабы могут менять окраску как в течение одной линьки, так и между линьками. Как и многим другим животным, это, по-видимому, позволяет им соответствовать внешнему виду фона для маскировки.
Рисунок 2. Береговые крабы ( Carcinus maenas ), содержащиеся на неоднородном светло-желтом песчаном фоне, могут, по-видимому, претерпевать значительные изменения в цвете и рисунке во время линьки (слева = до линьки, справа = после) .
Рисунок 3. Береговые крабы ( Carcinus maenas ) различаются в зависимости от среды обитания, в которой они обитают . Особи из относительно однородных коричнево-зеленых илистых отмелей (два столбца слева) более однородны и имеют сходный цвет с субстратом, тогда как особи из местообитаний красочных каменистых бассейнов (два столбца справа) гораздо более разнообразны и узорчаты.В каждом случае колонка крабов слева от пары — это взрослые особи, которые часто более однородны и менее изменчивы, чем молодые особи. Изображения изменены из Stevens et al. (2014б).
Рисунок 4. Значительные различия в разнообразии прибрежных крабов ( Carcinus maenas ) . Все эти особи (смесь взрослых и молодых особей) происходят только из одного каменного бассейна в Корнуолле, Великобритания.
Несмотря на прогресс в ряде областей, касающихся изменения окраски и маскировки животных, остается ряд важных нерешенных вопросов, касающихся не только крабов, но и целого ряда таксонов животных.Ниже я приведу шесть наиболее важных.
Какие животные меняют цвет для маскировки и что движет эволюцией изменения цвета?
Животные меняют цвет по нескольким причинам, не только для маскировки, но и для терморегуляции, подачи сигналов и защиты от ультрафиолетового (УФ) света. Было проведено несколько сравнительных анализов для полного изучения движущих сил изменения окраски у широко распространенных таксонов животных, и они были бы полезны для выявления задействованных экологических факторов (Stuart-Fox and Moussalli, 2009; Umbers et al. , 2014). Конечно, изменение цвета для маскировки было зарегистрировано у многих групп животных, включая чешуекрылых, ракообразных, головоногих, рептилий, амфибий, рыб и многих других. У видов Uca циркадные изменения могут обеспечить дневную защиту от ультрафиолетового излучения в более низких широтах. Однако защита от ультрафиолетовых лучей менее вероятна у прибрежных крабов с более умеренным климатом, особенно с учетом северного распространения их ареала. Напротив, причиной может быть терморегуляция (на мелководье или на открытом воздухе) в течение дня.У крабов-призраков, которые становятся светлее в течение дня, функция, по-видимому, заключается в маскировке, поскольку это изменение улучшает уровень соответствия пляжному субстрату. Для всех трех видов крабов кратковременные изменения в соответствии с фоновой яркостью, по-видимому, служат для маскировки. Обратите внимание, однако, что было проведено несколько прямых экспериментальных испытаний этих предложенных функций (см. Ниже).
Еще одна важная недостаточно изученная область — это то, как изменения внешнего вида действуют в различных временных масштабах и задействованных механизмах, начиная от кратковременного изменения цвета и заканчивая долгосрочной фенотипической пластичностью (Nettle and Bateson, 2015).Большая часть работ по изменению цвета проводилась на животных, которые быстро меняют цвет (таких как каракатицы и хамелеоны), или в основном была сосредоточена на кратковременных изменениях у таких видов, как береговые крабы. Как обсуждалось выше, многие из наиболее драматических изменений внешнего вида у многих видов, вероятно, связаны с линькой (заменой экзоскелета у беспозвоночных и заменой меха или перьев у позвоночных). Такие процессы редко проверялись или оценивались количественно, но они могут иметь большое значение для сопоставления с окружающей средой и факторами внутривидового разнообразия.
Одним из преимуществ фенотипической пластичности является то, что она позволяет животным изменять свой внешний вид в зависимости от среды обитания, в которой они живут, или даже напоминать определенные фоновые типы/микросреды обитания (Keeble and Gamble, 1899; Gamble and Keeble, 1900; Rosenblum, 2006; Todd et al. , 2006; Stevens et al., 2014b, 2015; Hultgren and Mittelstaedt, 2015; Jensen and Egnotovich, 2015; Russell and Dierssen, 2015; Duarte and Flores, 2016). Это может быть особенно ценным для видов с высокой степенью расселения и планктонных личиночных стадий, таких как многие ракообразные, поскольку может быть неопределенность в отношении того, где будет оседать молодь и какова визуальная среда.Еще одной особенностью многих из этих видов является то, что они имеют высокий уровень внутривидового разнообразия по окраске и рисунку (рис. 4, 5) даже в одной и той же местности (Keeble and Gamble, 1899; Gamble and Keeble, 1900; Todd et al., 2006). ; Stevens et al., 2014b; Carvalho-Batista et al., 2015; Hultgren and Mittelstaedt, 2015; Jensen and Egnotovich, 2015; Duarte and Flores, 2016). Причины этого пока во многом неясны. У таких видов, как береговой краб, это может позволить сопоставить ряд различных типов пятен в неоднородных средах обитания или предотвратить формирование изображений поиска хищника (Stevens et al. , 2014б). Они также могут представлять различные типы стратегий маскировки, такие как совпадение фона или разрушительная окраска. Еще один в значительной степени нерешенный вопрос заключается в том, почему многие крабы сильно различаются по внешнему виду среди особей в молодости, но претерпевают онтогенетические изменения по мере своего роста (рис. 3). У многих видов взрослые особи становятся менее узорчатыми, часто более темными и, по-видимому, менее загадочными (Palma and Steneck, 2001; Todd et al., 2006, 2009; Stevens et al., 2014b; Carvalho-Batista et al., 2015; Jensen and Эгнотович, 2015; Рассел и Дирсен, 2015).Два возможных объяснения заключаются в том, что более старые/более крупные крабы перемещаются в другие типы местообитаний, требуя другого типа маскировки, или что по мере того, как они становятся больше, они становятся менее восприимчивыми к хищникам (Todd et al., 2009). У береговых крабов молодь, вероятно, очень уязвима как для рыб, так и для хищников-птиц, тогда как взрослые особи более устойчивы к нападению со стороны мелких и средних рыб. Тем не менее функция этого паттерна онтогенетических изменений несколько загадочна. Это может помочь нам узнать, как маскировка работает и взаимодействует с размером тела, а также о компромиссах с другими функциями.
Рисунок 5. Многие виды крабов имеют чрезвычайно высокий уровень индивидуальной изменчивости в молодости, например, эти бороздчатые крабы ( Xantho incisus ) .
Как работает изменение цвета и пластичность?
За некоторыми заметными исключениями (например, хамелеоны Teyssier et al., 2015) способность многих животных изменять окраску в основном изучалась с точки зрения изменений хроматофорных клеток, содержащих различные пигменты (хотя в природе встречаются и другие механизмы Umbers et al., 2014). Изменения часто называют «физиологическими», когда они связаны с изменением состояния хроматофорных клеток и диспергированием пигмента в течение минут и часов, и «морфологическими», когда изменения происходят в течение дней и недель вследствие синтеза и перераспределения пигмента и клеток в организме. Стюарт-Фокс и Муссалли, 2009 г.). Опять же, у крабов была проделана большая работа по физиологическим изменениям.
Наблюдения под монокулярным микроскопом позволяют предположить, что у береговых крабов C. maenas имеет по крайней мере три типа хроматофоров с красным, белым и черным пигментом (Powell, 1962a).Они не были должным образом идентифицированы, но, вероятно, это эритрофоры (красный каротиноидный пигмент), лейкофоры или иридофоры (белые пурины) и меланофоры (черный/коричневый меланиновый пигмент). Береговые крабы также могут иметь ксантофоры (желтые птеридиновые клетки), которые могут позволить им стать более желтыми (рис. 2). Исследования Uca выявили сходные типы хроматофоров. Однако в прошлых работах по большинству видов обычно использовались субъективные показатели состояния хроматофора или качественные оценки в течение короткого промежутка времени (Fingerman and Yamamoto, 1967; Shibley, 1968; Jensen and Egnotovich, 2015), а не подробная гистология, окрашивание и идентификация пигмента в течение длительного времени. различные временные масштабы (особенно онтогенетические изменения) и не связывали это с количественной оценкой изменений общей окраски.Кроме того, то, что происходит во время линьки у ракообразных, насколько мне известно, не исследовано. В идеале будущая работа должна сочетать современные достижения в идентификации пигментных клеток и типов (например, Saenko et al., 2013) с количественной оценкой и визуальным моделированием изменений общего внешнего вида. Вдобавок ко всему, роль генетики и молекулярных путей в потенциале и функционировании изменения цвета пока неизвестна.
Также широко распространено мнение, что в дополнение к прямому действию света на хроматофорные клетки (первичная реакция) изменение цвета опосредовано зрительной информацией, полученной животным об окружающей среде, в которой оно живет (вторичная реакция; Umbers et al., 2014). В частности, часто предполагается, что ответы опосредованы не общим уровнем освещенности, а соотношением света, падающего на животное, и света, отраженного от субстрата под ним, что позволяет людям подбирать яркость фона независимо от общего уровня освещенности (Кибл). и Гэмбл, 1899 г.; Гэмбл и Кибл, 1900 г.; Самнер и Киз, 1929 г.). Это согласуется с работой, показывающей, что, например, крабы-призраки не просто становятся темнее, если их поместить в темноту (Stevens et al., 2013). Такая информация может быть получена при различном излучении света, падающего на разные части глаза (Браун и Сандин, 1948). Что касается цвета, у животных может быть либо система цветового зрения, способная различать разные цвета фона, либо, как в случае с каракатицей, они могут быть дальтониками и потенциально использовать эмпирическое правило о том, как различная яркость и объекты преобразуются в цвет. , в сочетании с цветными пигментами, близкими к природным субстратам (Mäthger et al., 2006, 2008). В качестве альтернативы или в дополнение каракатица может в первую очередь соответствовать яркости и рисунку фона, если это наиболее важно для облегчения маскировки. Хотя эти идеи кажутся логичными и интуитивными, очень немногие эксперименты проверяли эти предположения. Наша работа с береговыми крабами (Stevens et al. , 2014a) не выявила признаков изменения цвета на красном и зеленом фоне. Это может произойти, если крабы не могут определить разницу между этими цветами (или при отсутствии различий в яркости, если крабы используют ахроматические механизмы для сопоставления цветов).Кажется вероятным, что береговые крабы могут обнаруживать различия между относительно коротковолновым («синим») и длинноволновым («красным» или «желтым») светом (Martin and Mote, 1982), но неясно, могут ли они различать длинноволновый и средневолновой свет. («зеленый», «красный» и «желтый»). Однако более долгосрочные изменения цвета все же происходят (рис. 2), поэтому, скорее всего, проблема в том, что временной масштаб наших экспериментов был очень коротким.
Дальнейшие исследования проверили, как дисперсия пигмента в хроматофорных клетках может быть связана с визуальной информацией.У Uca и других ракообразных состояние экспансии хроматофорных клеток по крайней мере частично обусловлено гормонами, выделяемыми стебельками глаз (например, Fingerman and Yamamoto, 1967; Fingerman, 1973). Однако исследования крабов Рак с удалением либо целых стебельков глаз, либо только комплекса синусовых желез (источник гормонов, диспергирующих пигмент, на стебельке глаз), либо разрезания зрительных нервов позволяют предположить, что визуальная информация через зрительный нерв является ключом к контролю дисперсии пигмента, а не наличие только синусовых желез (Shibley, 1968).То есть визуальные сигналы, проходящие к зрительной доле, кажутся решающими в контроле ответов (предположительно, через другие источники гормонов, не связанные с стебельками глаз). Поэтому необходимо проверить, как визуальная информация и роль гормонов связаны у крабов и других животных, чтобы понять пути, которыми контролируются изменения.
Дополнительное недавнее соображение заключается в том, что изменение цвета у некоторых видов может управляться фоторецепторами, которые находятся вне глаз. Например, кожа некоторых головоногих содержит белки опсины, которые используются для обнаружения света (Mäthger et al. , 2010). Информация от этих белков может использоваться для дополнения изменения цвета, управляемого глазами, и даже позволяет частям тела настраивать свое соответствие окружающей среде. Это может возникать за счет сравнения с информацией от хроматофорных клеток, также чувствительных к свету (Kingston et al., 2015; Ramirez, Oakley, 2015). Другая недавняя работа показала, что некоторые гекконы могут менять цвет, чтобы соответствовать фону, когда их глаза закрыты, но не когда закрыты их бока (которые, по-видимому, содержат опсины; Fulgione et al., 2014). Необходима дальнейшая работа над ролью обнаружения экстраокулярного света в управлении изменением цвета.
Изменение цвета также оказалось ценной системой для понимания зрения и восприятия. Управляя фоном контролируемым образом, исследователи смогли определить, на какие сигналы реагируют такие животные, как головоногие моллюски, и почему, основываясь на том, как они меняют узоры. Это показало, что, среди прочего, головоногие моллюски оценивают аспекты интенсивности фона, кодируют местоположение визуальных краев, воспринимают сигналы глубины и даже выполняют завершение контуров, восполняя недостающую информацию из визуальной сцены (Kelman et al. , 2007; Барбоза и др., 2008 г.; Жилинский и др., 2009, 2012, 2016).
Наконец, мы должны отметить, что изменение цвета не всегда зависит исключительно от визуальной информации. У некоторых креветок и гусениц ранние и недавние исследования свидетельствуют о том, что диета может играть важную роль, отдельно или в сочетании со зрением, во влиянии на изменения внешнего вида (Keeble and Gamble, 1899; Gamble and Keeble, 1900; Greene, 1996; Noor). и др., 2008).
Какую функцию выполняют циркадные ритмы изменения цвета?
Для многих видов характерна смена дня и ночи.Например, помимо крабов, креветки-хамелеоны ( Hippolyte varians ) меняют цвет на сине-серый, и животные из различных других таксонов также проходят циклы (Gamble and Keeble, 1900; Powell, 1962a; Darnell, 2012; Stevens et al. ., 2013). Функции этих циклов, насколько мне известно, редко проверялись. Как обсуждалось выше, у крабов-призраков это может способствовать маскировке, а у креветок-хамелеонов это может способствовать лучшему приспособлению к условиям лунного света. Однако у всех видов часто предполагается, но, по-видимому, никогда не проверялось, что изменение окраски ночью также может быть механизмом экономии энергии.
Сколько стоит изменение цвета?
Широко распространено мнение, что изменение цвета связано с энергетическими/метаболическими затратами, по крайней мере, с точки зрения изменения состояния хроматофорных клеток и синтеза нового пигмента и клеток, однако до сих пор это никогда не проверялось (Stuart-Fox and Moussalli, 2009). Если процесс расширения или сокращения хроматофорных клеток требует энергии, то изменения в ночное время могут быть механизмом экономии затрат, например, за счет расслабления мышц и изменений в клеточном состоянии, когда снижается зрительное хищничество и потребность в маскировке.Ночью давление со стороны визуальных хищников, вероятно, уменьшается (хотя это предположение требует проверки), или крабы также могут больше прятаться под камнями и зарываться в субстрат. Наша работа как с крабами-призраками, так и с береговыми крабами выявила значительные различия в способности изменять цвет/яркость (Stevens et al. , 2013, 2014a). Это может быть связано с модификацией ритмов изменения окраски, связанных с другими факторами, такими как приливы и лунные циклы, как это имеет место у некоторых видов Uca (Fingerman, 1956; Fingerman et al., 1958), но в равной степени могут отражать различия в способности некоторых людей изменять цвет в зависимости от их состояния. Другие затраты и ограничения также могут быть связаны с изменением цвета, включая конкурирующие требования, такие как камуфляж, терморегуляция и защита от ультрафиолета.
Каково адаптивное значение изменения цвета и пластичности?
Возможно, самый очевидный пробел в нашем понимании камуфляжа и изменения цвета заключается в том, как это напрямую влияет на риск нападения хищников; то есть адаптивное значение, которое обеспечивает изменение цвета для маскировки.Немногие исследования, если вообще проводились, проверяли, как изменение цвета влияет на поведение хищников и обнаружение хищниками, или делали это косвенно (например, Hultgren and Mittelstaedt, 2015), и это должно стать главной целью на будущее. В некотором отношении это отсутствие доказательств неудивительно. Самая последняя работа по изучению эффективности маскировки проводилась в искусственных системах с искусственными раздражителями (например, Bond and Kamil, 2002; Cuthill et al., 2005; Merilaita and Lind, 2005; Webster et al., 2013) и даже в В дикой природе только недавно исследования показали, что уровень маскировки животного напрямую влияет на вероятность нападения (Troscianko et al., 2016). Тем не менее, необходимы контролируемые эксперименты по проверке того, как изменение цвета влияет на риск нападения хищников.
Позволит ли изменение цвета некоторым животным справиться с последствиями меняющегося мира?
Логически можно было бы ожидать, что способность изменять внешний вид может помочь некоторым видам приспособиться к изменяющемуся миру перед лицом вмешательства человека. Например, если в визуальной среде происходят изменения, изменение цвета может позволить камуфляжу оставаться эффективным. Однако это, вероятно, будет сильно зависеть от механизмов, управляющих изменениями. Например, у зайцев-снегоступов уменьшение снежного покрова и продолжительности из-за изменения климата меняет внешний вид фона, но до сих пор нет доказательств того, что зайцы могут изменять время линьки для перехода от коричневого к белому меху и обратно в соответствии со снегом. смены стаи (Зимова и др., 2014). В результате люди потенциально станут более несовместимыми с окружающей средой, что, в свою очередь, может нарушить их камуфляж и тем самым увеличить вероятность хищничества. Однако это сезонные изменения, тогда как изменения внешнего вида, происходящие в более короткие промежутки времени, могут в большей степени способствовать преодолению изменений, вызванных деятельностью человека.Световое загрязнение также является серьезной проблемой, затрагивающей большую часть мира, и может изменить визуальную среду, особенно если спектр света в ночное время отличается от спектра лунного и солнечного света (Gaston et al., 2013). Остается без ответа вопрос, в какой степени изменение цвета может помочь животным справиться с этим. Потенциально, если спектр искусственного света сильно отличается от дневного света, то животные, такие как крабы, могут менять цвет ночью в неестественно ярких условиях на внешний вид, который не соответствует фону при естественном освещении в течение дня, а это означает, что изменение цвета ночью фактически уменьшается. дневной камуфляж.Еще одним соображением является температура и кислотность океана, связанные с изменением климата. Имеются данные о том, что изменение цвета крабов может зависеть от температуры (например, Powell, 1962b), а недавняя работа показала, как подкисление океана может влиять на поведенческие и физиологические реакции личинок рыб (Munday et al., 2009). Как изменения климата и океанов повлияют на способность животных менять цвет, еще неизвестно.
Изменение цвета и маскировка оказались ценной системой для изучения визуального восприятия, физиологии и поведения.Однако многие ключевые вопросы остаются без ответа или непроверенными, и этот предмет может многое рассказать об эволюции, внутривидовом разнообразии и адаптации к меняющимся условиям. В конечном счете, этот предмет должен оставаться прекрасным примером того, как исследования могут сочетать ряд междисциплинарных методов, от клеточных исследований до моделирования зрения и поведенческих экспериментов, и решать проблемы, начиная от эволюционно-экологических вопросов «сверху вниз» и заканчивая «сверху вниз». механистические вопросы «снизу вверх» (Kemp et al., 2015).
Вклад авторов
Автор подтверждает, что является единственным автором данной работы и одобрил ее публикацию.
Финансирование
MS финансировался Исследовательским советом по биотехнологии и биологическим наукам Дэвида Филлипса (BB/G022887/1).
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Я очень благодарен двум рецензентам и многочисленным коллегам за обсуждение и сотрудничество, в том числе Питеру Тодду, Дженни Кумс, Дженни Изли, Осси Нокелайнен, Наташе Прайс и Саре Майнотт.
Биография автора
Мартин Стивенс — адъюнкт-профессор сенсорной и эволюционной экологии Эксетерского университета. Он защитил докторскую диссертацию в Бристольском университете по зрению птиц и маскировке животных, а затем прошел несколько стипендий в Кембриджском университете, прежде чем переехать в Эксетер в 2013 году.Его работа была сосредоточена на окраске животных, зрении и методах количественной оценки визуальных сигналов, особенно на анализе изображений. Это включало работу над камуфляжем, мимикрией, выводковыми паразитами и сексуальными сигналами. Основная текущая область работы — понять адаптивную ценность и механизмы изменения цвета и маскировки.
Ссылки
Абрамовиц, А.А. (1937). Хроматофоротропный гормон ракообразных: стандартизация, свойства и физиология желез глазного стебля. Биол. Бык. 72, 344–365. дои: 10.2307/1537694
Полнотекстовая перекрестная ссылка | Академия Google
Аткинс, Д. (1926). О ночном изменении окраски краба-горошки ( Pinnotheres veterum ). Природа 117, 415–416. дои: 10.1038/117415b0
Полнотекстовая перекрестная ссылка | Академия Google
Барбоза, А., Метгер, Л.М., Буреш, К.С., Келли, Дж., Чабб, К.-К., и Хэнлон, Р.Т. (2008). Камуфляж каракатицы: влияние контраста и размера субстрата на создание однородных, пятнистых или разрушительных рисунков тела. Виз. Рез. 48, 1242–1253. doi: 10.1016/j.visres.2008.02.011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Браун, Ф.А., и Сандин, М.И. (1948). Реакция хроматофоров краба-скрипача Uca на свет и температуру. Физиол. Зоопарк. 21, 361–371. doi: 10.1086/physzool.21.4.30152015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Браун, Ф. А., и Уэбб, Х. М. (1948). Температурные отношения эндогенной суточной ритмичности краба-скрипача, Uca . Физиол. Зоопарк. 21, 371–381. doi: 10.1086/physzool.21.4.30152016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Карвальо-Батиста, А., Песчинелли, Р. А., Гарсия, Дж. Р., Герра, П. Г. М., Пардо, Л. М., и Мантелатто, Ф. Л. (2015). Crypsis у грязевого краба Panopeus americanus Saussure, 1857 (Decapoda, Panopeidae): связь с половой зрелостью. Ракообразная 88, 963–977. дои: 10.1163/15685403-00003461
Полнотекстовая перекрестная ссылка | Академия Google
Цзяо, К.К., Викизер, Дж. К., Аллен, Дж. Дж., Гентер, Б., и Хэнлон, Р. Т. (2011). Гиперспектральная визуализация камуфляжа каракатиц указывает на хорошее совпадение цветов в глазах рыб-хищников. Проц. Натл. акад. науч. США 108, 9148–9153. doi: 10.1073/pnas.10190
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кронин, Т.В.-Т., Чиоу, Х., Колдуэлл, Р.Л.Р.Н., и Маршалл, Дж. (2009). Сигналы поляризации у креветок-богомолов. Проц. SPIE 7461, 74610. дои: 10.1117/12.828492
Полнотекстовая перекрестная ссылка | Академия Google
Катхилл, И. К., Стивенс, М., Шеппард, Дж., Мэддокс, Т., Паррага, К. А., и Трощанко, Т. С. (2005). Подрывная окраска и сопоставление фонового рисунка. Природа 434, 72–74. doi: 10.1038/nature03312
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дарлинг, Дж. А., Бэгли, М. Дж., Роман, Дж., Теполт, С. К., и Геллер, Дж. Б. (2008). Генетические паттерны нескольких интродукций глобально инвазивного рода крабов Carcinus . Молекуляр. Экол. 17, 4992–5007. doi: 10.1111/j.1365-294X.2008.03978.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дарнелл, М. З. (2012). Экологическая физиология циркадного ритма пигментации у краба-скрипача Uca panacea . Дж. Экспл. Морская биол. Экол. 426–427, 39–47. doi: 10.1016/j.jembe.2012.05.014
Полнотекстовая перекрестная ссылка | Академия Google
Детто Т. , Хемми Дж. М. и Бэквелл П. Р. Ю. (2008).Изменение окраски и цвета краба-скрипача, Uca capricornis : описательное исследование. PLoS ONE 3:e1629. doi: 10.1371/journal.pone.0001629
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дуарте, Р. К., и Флорес, А. В. (2016). Морфо-специфическая среда обитания и распределение по полу у креветок-каридов Hippolyte obliquimanus . J. Marine Biol. Доцент . дои: 10.1017/s0025315416000230. [Epub перед печатью].
Полнотекстовая перекрестная ссылка
Фингерман, М.(1955). Стойкие суточные и приливные ритмы изменения окраски у Callinectes sapidus . Биол. Пуля. 109, 255–264. дои: 10.2307/1538725
Полнотекстовая перекрестная ссылка | Академия Google
Фингерман, М. (1956). Различие в приливных ритмах изменения окраски двух видов крабов-скрипачей. Биол. Пуля. 110, 274–290. дои: 10.2307/1538833
Полнотекстовая перекрестная ссылка | Академия Google
Фингерман, М. (1973). Поведение хроматофоров краба-скрипача Uca pugilator и карликового рака Cambarellus shufeldti в ответ на синтетический Pandalus красный пигмент-концентрирующий гормон. Ген. комп. Эндокрин. 20, 589–592. дои: 10.1016/0016-6480(73)
-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фингерман, М., Лоу, М.Е., и Мобберли, В.К. (1958). Факторы окружающей среды, участвующие в установлении фаз приливного ритма изменения окраски у крабов-скрипачей Uca pugilator и Uca minax . Лимнол. Океаног. 3, 271–282. doi: 10.4319/lo.1958.3.3.0271
Полнотекстовая перекрестная ссылка | Академия Google
Фингерман, М.и Ямамото, Ю. (1967). Суточный ритм миграции меланофорного пигмента у бесглазных крабов-скрипачей, Uca pugilator (Bosc). Ракообразные 12, 303–319. дои: 10.1163/156854067X00279
Полнотекстовая перекрестная ссылка | Академия Google
Фульджионе Д. , Трапанезе М., Маселли В., Риппа Д., Итри Ф., Аваллоне Б. и Райя П. (2014). Видение сквозь кожу: светочувствительность кожи обеспечивает криптизм у мавританского геккона. Дж. Зул. 294, 122–128. дои: 10.1111/jzo.12159
Полнотекстовая перекрестная ссылка | Академия Google
Гэмбл, Ф.В., и Кибл, Ф.В. (1900). Варианты Ипполита : исследование изменения цвета. J. Cell Sci. С2-43, 589–698.
Академия Google
Гастон, К. Дж., Бенни, Дж., Дэвис, Т. В., и Хопкинс, Дж. (2013). Экологические последствия ночного светового загрязнения: механистическая оценка. Биол. Ред. 88, 912–927. doi: 10.1111/brv.12036
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Грин, Э.(1996). Влияние качества света и рациона личинок на индукцию морфа у полиморфной гусеницы Nemoria arizonaria (Lepidoptera: Geometridae). Биол. Дж. Линн. соц. 58, 277–285. doi: 10.1111/j.1095-8312.1996.tb01435.x
Полнотекстовая перекрестная ссылка | Академия Google
Hanlon, R. T.-C., Chiao, C., Mäthger, L.M., Barbosa, A., Buresch, K.C., и Chubb, C. (2009). Динамический камуфляж головоногих: преодоление континуума между совпадением фона и разрушительной окраской. Фил. Транс. Р. Соц. В 364, 429–437. doi: 10.1098/rstb.2008.0270
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хемми, Дж. М., Маршалл, Дж., Пикс, В., Воробьев, М., и Зейл, Дж. (2006). Переменные цвета краба-скрипача Uca vomeris и их связь с фоном и хищничеством. Дж. Экспл. биол. 209, 4140–4153. doi: 10.1242/jeb.02483
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Полнотекстовая перекрестная ссылка | Академия Google
Халтгрен, К.М., и Миттельштадт, Х. (2015). Изменение цвета морской изопод является адаптивным для уменьшения хищничества. Курс. Зоол. 61, 739–748. doi: 10.1093/czoolo/61.4.739
Полнотекстовая перекрестная ссылка | Академия Google
Дженсен, Г. К., и Эгнотович, М. С.(2015). Более белый оттенок самца: совпадение цветового фона в зависимости от размера и пола у желтого берегового краба Hemigrapsus oregonensis (Dana, 1851). Курс. Зоол. 61, 729–738. doi: 10.1093/czoolo/61.4.729
Полнотекстовая перекрестная ссылка | Академия Google
Кибл, Ф.В., и Гэмбл, Ф.В. (1899 г.). Цветофизиология Ипполита вариантов . Проц. Р. Соц. Б 65, 461–468. doi: 10.1098/rspl.1899.0059
Полнотекстовая перекрестная ссылка
Кельман, Э., Баддели Р., Шохет А. и Осорио Д. (2007). Восприятие визуальной текстуры и проявление разрушительного камуфляжа каракатицей, Sepia officinalis . Проц. Р. Соц. В 274, 1369–1375. doi: 10.1098/rspb.2007.0240
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кемп, Д. Дж., Герберштейн, М. Е., Флейшман, Л. Дж., Эндлер, Дж. А., Беннетт, А. Т. Д., и Уайтинг, М. Дж. (2015). Интегративная основа для оценки окраски в природе. утра. Нац. 185, 705–724. дои: 10.1086/681021
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кингстон, А.С.Н., Кузириан, А.М., Хэнлон, Р.Т., и Кронин, Т.В. (2015). Компоненты зрительной фототрансдукции в хроматофорах головоногих предполагают кожную фоторецепцию. Дж. Экспл. биол. 218, 1596–1602. doi: 10.1242/jeb.117945
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лигон, Р. А., и Макгроу, К. Дж. (2013).Хамелеоны общаются сложными изменениями цвета во время состязаний: разные участки тела передают разную информацию. Биол. лат. 9, 20130892. doi: 10.1098/rsbl.2013.0892
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мартин, Ф.Г., и Моут, М.И. (1982). Цветовые рецепторы морских ракообразных: второй спектральный класс ретинулярных клеток в сложных глазах Callinectes и Carcinus . J. Комп. Физиол. А 145, 549–554.дои: 10.1007/BF00612820
Полнотекстовая перекрестная ссылка | Академия Google
Метгер, Л. М., Барбоза, А., Майнер, С., и Хэнлон, Р. Т. (2006). Дальтонизм и контрастное восприятие у каракатиц ( Sepia officinalis ), определяемые с помощью визуального сенсомоторного анализа. Виз. Рез. 46, 1746–1753 гг. doi: 10.1016/j.visres.2005.09.035
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Метгер, Л. М.-К., Чиао, К., Барбоза, А., и Хэнлон, Р.Т. (2008). Подбор цвета на естественных субстратах у каракатиц, Sepia officinalis . J. Комп. Физиол. А 194, 577–585. doi: 10.1007/s00359-008-0332-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Метгер, Л. М., Робертс, С. Б., и Хэнлон, Р. Т. (2010). Доказательства распределенной светочувствительности кожи каракатиц, Sepia officinalis . Биол. лат. 6, 600–603. doi: 10.1098/rsbl. 2010.0223
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Макгоу, И.Дж., Эджелл, Т.С., и Кайзер, М.Дж. (2011). Демография популяций местных и недавно инвазированных популяций зеленого краба Carcinus maenas . Морской экол. прог. сер. 430, 235–240. дои: 10.3354/meps09037
Полнотекстовая перекрестная ссылка | Академия Google
Мерилаита, С., и Линд, Дж. (2005). Совпадающая с фоном и разрушительная окраска, а также эволюция загадочной окраски. Проц. Р. Соц. В 272, 665–670. doi: 10.1098/rspb.2004.3000
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мандей, П.Л., Диксон, Д.Л., Донельсон, Дж.М., Джонс, Г.П., Пратчетт, М.С., и Дёвинг, К.Б. (2009). Закисление океана ухудшает обонятельную дискриминацию и способность морской рыбы к самонаведению. Проц. Натл. акад. науч. США 106, 1848–1852 гг. doi: 10.1073/pnas.0809996106
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нахман М. В., Хекстра Х.Е. и Д’Агостино С.Л. (2003). Генетическая основа адаптивного меланизма у карманных мышей. Проц. Натл. акад. науч.США 100, 5268–5273. doi: 10.1073/pnas.0431157100
Реферат PubMed | Полный текст перекрестной ссылки
Крапива, Д., и Бейтсон, М. (2015). Адаптивная пластичность развития: что это такое, как мы можем ее распознать и когда она может развиться? Проц. Р. Соц. B 282, 20151005. doi: 10.1098/rspb.2015.1005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нур, М.А.Ф., Парнелл, Р.С., и Грант, Б.С. (2008). Обратимый полифенизм окраски гусениц американской перечной огневки ( Biston betularia cognataria ). PLoS ONE 3:e3142. doi: 10.1371/journal.pone.0003142
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пальма, А.Т., и Стенек, Р.С. (2001). Снижает ли переменная окраска молодых морских крабов риск визуального хищничества? Экология 82, 2961–2967. doi: 10.1890/0012-9658(2001)082[2961:DVCIJM]2.0.CO;2
Полнотекстовая перекрестная ссылка | Академия Google
Пауэлл, Б.Л. (1962a). Распределение и ритмическое поведение хроматофоров молоди Carcinus maenas (L.). Дж. Аним. Экол 31, 251–261. дои: 10.2307/2139
Полнотекстовая перекрестная ссылка | Академия Google
Пауэлл, Б.Л. (1962b). Реакция хроматофоров Carcinus maenas (L. 1758) на свет и температуру. Ракообразные 4, 93–102. дои: 10.1163/156854062X00120
Полнотекстовая перекрестная ссылка | Академия Google
Рамирес, доктор медицины, и Окли, Т. Х. (2015). Независимое от глаза, активируемое светом расширение хроматофора (LACE) и экспрессия генов фототрансдукции в коже Octopus bimaculoides . Дж. Экспл. биол. 218, 1513–1520. doi: 10.1242/jeb.110908
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рассел, Б.Дж., и Дирсен, Х.М. (2015). Использование гиперспектральных изображений для оценки загадочного соответствия цветов у саргассовых крабов, связанных с . PLoS ONE 10:e0136260. doi: 10.1371/journal.pone.0136260
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Саенко С.В., Тейссье Дж., ван дер Марель Д.и Милинкович, MC (2013). Точная колокализация взаимодействующих структурных и пигментных элементов приводит к обширным вариациям цветового рисунка у ящериц Phelsuma . БМС Биол. 11:105. дои: 10.1186/1741-7007-11-105
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стивенс, М., Бродерик, А.С., Годли, Б.Дж., Лаун, А.Е., Трощанко, Дж., и Вебер, С.Б. (2015). Соответствие фенотипа и среды у песчаных блох. Биол. лат. 11, 20150494.doi: 10.1098/rsbl.2015.0494
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стивенс, М., Лаун, А. Э., и Вуд, Л. Э. (2014a). Изменение окраски и маскировка у молоди береговых крабов Carcinus maenas . Перед. Экол. Эвол. 2:14. doi: 10.3389/fevo.2014.00014
Полнотекстовая перекрестная ссылка | Академия Google
Стивенс, М. , Пей Ронг, К., и Тодд, П. А. (2013). Изменение цвета и маскировка у рогатого краба-призрака Ocypode ceratophthalmus . Биол. Дж. Линн. соц. 109, 257–270. doi: 10.1111/bij.12039
Полнотекстовая перекрестная ссылка | Академия Google
Стивенс М., Вуд Л. Э. и Лаун А. Э. (2014b). Камуфляж и индивидуальная изменчивость береговых крабов ( Carcinus maenas ) из разных местообитаний. PLoS ONE 9:e115586. doi: 10.1371/journal.pone.0115586
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стюарт-Фокс, Д., и Муссалли, А. (2009). Камуфляж, общение и терморегуляция: уроки организмов, меняющих цвет. Филос. Транс. Р. Соц. В 364, 463–470. doi: 10.1098/rstb.2008.0254
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Самнер, Ф. Б., и Киз, А. Б. (1929). Влияние различий в видимом источнике освещения на тень, которую принимает камбала на данном фоне. Физиол. Зоол. 2, 495–504. doi: 10.1086/physzool.2.4.30152972
Полнотекстовая перекрестная ссылка | Академия Google
Тейссье Дж., Саенко С.В., ван дер Марел Д.и Милинкович, MC (2015). Фотонные кристаллы вызывают активное изменение цвета у хамелеонов. Нац. Комм. 6, 6368. doi: 10.1038/ncomms7368
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тодд П.А., Брайерс Р.А., Лэдл Р.Дж. и Миддлтон Ф. (2006). Совпадение фенотипа и среды у берегового краба ( Carcinus maenas ). Морская биология. 148, 1357–1367. doi: 10.1007/s00227-005-0159-2
Полнотекстовая перекрестная ссылка | Академия Google
Тодд, П.А., О, Дж., Локе, Л.Х.Л., и Лэдл, Р.Дж. (2012). Многомасштабное совпадение фенотипа и субстрата: данные береговых крабов ( Carcinus maenas L.). Экол. Сложный. 12, 58–62. doi: 10.1016/j.ecocom.2012.09.005
Полнотекстовая перекрестная ссылка | Академия Google
Тодд П.А., Цю В. и Чонг К.Ю. (2009). Онтогенетические сдвиги в строении панциря и/или окраске литоральных и сублиторальных брахиурановых крабов. Зоопарк Raffles Bullet. 57, 543–550.
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зимова, М., Миллс, Л.С., Лукач, П.М., и Митчелл, М.С. (2014). Зайцы-снегоступы демонстрируют ограниченную фенотипическую пластичность, чтобы не соответствовать сезонному камуфляжу. Проц. Р. Соц. B 281, 20140029. doi: 10.1098/rspb.2014.0029
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зилински, С.-А., Дармейлак, С., и Шашар, Н. (2012). Визуальная интерполяция для завершения контура европейской каракатицы ( Sepia officinalis ) и ее использование в динамическом камуфляже. Проц. Р. Соц. В 279, 2386–2390. doi: 10.1098/rspb.2012.0026
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зилински, С., и Йонсен, С. (2011). Мезопелагические головоногие моллюски переключаются между прозрачностью и пигментацией, чтобы оптимизировать маскировку на глубине. Курс. биол. 21, 1937–1941. doi: 10.1016/j.cub.2011.10.014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зилински С., Осорио Д. и Джонсен С. (2016). Каракатицы видят форму по затенению, точной настройке окраски в ответ на визуальные признаки глубины и направленное освещение. Проц. Р. Соц. B 283, 20160062. doi: 10.1098/rspb.2016.0062
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Жилинский С., Осорио Д.и Шохет, А. (2009). Восприятие краев и визуальной текстуры в камуфляже обыкновенной каракатицы, Sepia officinalis . Фил. Транс Р. Соц. В 364, 439–448. doi: 10. 1098/rstb.2008.0264
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Определение фенотипа и примеры — Биологический онлайн-словарь генетических особенностей и влияния окружающей среды.Фенотип организма включает его морфологические, биохимические, физиологические и поведенческие свойства. Таким образом, фенотип представляет собой совокупность характеристик, проявляемых организмом, которые являются результатом экспрессии генов организма, а также влияния факторов окружающей среды и случайных вариаций. Взаимодействие между этими факторами часто представлялось следующим соотношением: генотип + окружающая среда + случайная изменчивость → фенотип.
Давайте поговорим о фенотипе! Присоединяйтесь к обсуждению на нашем форуме: Неполное доминирование vs.кодоминирование. Станьте частью нашего сообщества!
Определение фенотипа
В биологии « фенотип » определяется как наблюдаемые и измеримые характеристики организма в результате взаимодействия генов организма, факторов окружающей среды и случайных вариаций . Фенотип организма будет включать не только наблюдаемые особенности, такие как морфология, но также будет включать молекулы и структуры, такие как РНК и белки, продуцируемые генами; это называется «молекулярным фенотипом».
Этимология
Термин фенотип поступил из латинской Phaeno- , от греческого Phaeno- , что означает «сияющий», от Файнейна, , что означает «Чтобы сиять» , «Чтобы появиться , « показать» , а — набрать из «опечатки». Сравните: генотип. Термин фенотипический представляет собой описательное слово, которое используется для обозначения, отношения или описания фенотипа конкретного организма.
Рисунок 1: Эта диаграмма (квадрат Пеннета) показывает взаимосвязь между фенотипом и генотипом. Характер окраски лепестков растений гороха представлен генами В и В . Ген B является доминантным признаком и проявляется в виде цветка с пурпурными лепестками. Ген b является рецессивным признаком. Тестовое скрещивание двух растений, гетерозиготных (Bb) по признаку пурпурной окраски лепестков, даст 3 потомства с признаком пурпурных цветков (BB и Bb) и 1 потомство с признаком белых цветков ( бб). В этом примере соотношение фенотипов составляет 3:1.
Признак против фенотипа
Признак — это атрибут фенотипа организма. Таким образом, в генетике этот признак также упоминается как фенотипический признак , чтобы отличить один признак от другого в более широком термине фенотип. Фенотип организма состоит из различных признаков. Признаки могут быть унаследованы (генетически детерминированы), приобретены под влиянием окружающей среды или в результате их взаимодействия.Например, цвет волос — это персонаж, черты которого могут быть черными, светлыми, рыжими или брюнетками.
Фенотип против генотипа
В генетике генотип и фенотип являются общими терминами, используемыми для объяснения внешнего вида, функций и поведения организма. Генотип определяется как набор генов, экспрессия которых определяет характеристику или признак организма. Проще говоря, генотип — это генетический вклад в фенотип.
Гены составляют последовательности ДНК. У людей и других организмов они встречаются парами — один исходит от родителя-мужчины, а другой — от родителя-женщины. Пары генов, занимающие одни и те же локусы на хромосомах и контролирующие один и тот же признак, называются аллелями. Итак, как связаны аллели и признаки?
Как правило, пара генов (или пара аллелей) определенного признака состоит из двух генов, один из которых доминантный, , а другой рецессивный. Доминантный аллель будет выражен как признак, тогда как рецессивный аллель не будет выражен. Аннотируя доминантный аллель A и рецессивный аллель a , можно выделить три возможных генотипа: (1) AA, гомозиготный доминантный аллель, (2) Aa, гетерозиготный доминантный и (3) ) а. о., гомозиготный рецессивный. Генотип организма является основной детерминантой фенотипа организма.
Но как генотип влияет на фенотип? Возьмем, к примеру, пару аллелей (или пару генов), определяющую конкретный признак, где один из них является доминантным ( А ), а другой рецессивным ( А ).Доминантный аллель ( А ) будет выражен и станет частью фенотипа организма, тогда как эффект рецессивного аллеля ( А ) будет замаскирован.
Когда признак следует менделевскому наследованию, A проявится как признак, а a — нет. Таким образом, фенотип организма должен включать признаки всех экспрессируемых генов. Однако у людей многие из наблюдаемых признаков более сложны, чем те, которые соответствуют менделевскому образцу.Сложные признаки, такие как рост и цвет кожи, возникают в результате взаимодействия нескольких аллелей, как в случае полигенного наследования.
Давайте изучим различные черты фенотипа! Присоединяйтесь к обсуждению на форуме: Неполное доминирование против кодоминирования.
Экспрессия
Фенотип является результатом генетических факторов, влияния окружающей среды и случайных генетических вариаций. Когда пара аллелей, определяющих тот или иной признак, состоит из одних и тех же генов, т.е. AA или аа, признак описан как гомозиготный. Когда аллельный состав состоит из разных генов, т.е. Аа, признак описан как гетерозиготный. Наличие доминантного аллеля, т.е. АА или Аа, приведет к проявлению признака ( А ), тогда как отсутствие доминантного аллеля, т.е.е. а.о., приведут к выражению другого признака ( a ). Это случай полного доминирования, и он соответствует менделевскому наследованию.
В случаях кодоминантности, неполного доминирования и полигенного наследования проявление признака не будет соответствовать менделевскому образцу. Например, при кодоминантности аллели пары генов в гетерозиготе будут полностью выражены, поскольку оба аллеля в паре являются доминантными над (например, AB). При неполном доминировании результирующий признак будет промежуточным эффектом двух аллелей. Это связано с тем, что доминантный аллель будет выражен лишь частично. В результате гетерозиготное потомство будет демонстрировать фенотип, промежуточный по отношению к фенотипу родителей.
Помимо генетических взаимодействий, фенотип организма будет зависеть также от влияния окружающей среды и случайных (генетических) вариаций. Воздействие факторов окружающей среды может повлиять на внешний вид организма. Например, светлая кожа, которая постоянно подвергается воздействию солнечных лучей, приводит к потемнению кожи из-за повышенного производства меланина.
Что касается случайных вариаций, то генетическая мутация может изменить физические признаки или, по крайней мере, приспособленность организма. Изменения в генах необходимы, поскольку они управляют эволюцией и естественным отбором. Генотипы, влияние окружающей среды и генетические вариации помогают объяснить, почему люди склонны иметь разные фенотипы.
Экстремальный фенотип
Экстремальный фенотип возникает, когда аллели родителей объединяются и приводят к гибриду с фенотипом, который на выше или на выше , чем фенотипы его родителей. Его трансгрессивный фенотип может быть полезным или вредным в зависимости от того, как он влияет на общую приспособленность потомства. Формирование крайних фенотипов называется трансгрессивной сегрегацией . Примером гибрида с экстремальным фенотипом является потомство от скрещивания Helianthus annuus и Helianthus petiolaris . Два вида подсолнечника дали гибриды с трансгрессивными признаками. Гибриды, в отличие от их родителей, способны процветать в областях, где их родители не могут.Они способны выживать в песчаных дюнах и солончаках. (Ref.1)
Рисунок 2: Пример экстремального фенотипа. Источник изображения: Мария Виктория Гонзага из Biology Online, из работ Fir0002 (обычная фотография подсолнуха — CC BY-SA 3.0), Министерства сельского хозяйства США (фото подсолнуха в прериях — общественное достояние) и Кенрайза ( Helianthus anomalus photo) .
Рекомбинантный фенотип
Мейоз является одним из важных биологических событий, которые приводят к большей изменчивости фенотипов организмов. В частности, во время метафазы мейоза I гомологичные хромосомы объединяются для обмена генами посредством гомологичной рекомбинации . Когда гомологичные хромосомы достигают конца мейоза (телофаза II), четыре дочерние клетки будут иметь хромосомы, отличные друг от друга. Некоторые из них станут гаметами, содержащими рекомбинантные гены. Когда такая гамета оплодотворяется, например, диким типом, она разовьется в потомство с рекомбинантным фенотипом, т.е.е. фенотип, отличный от фенотипов своих родителей.
Как определить фенотип рекомбинантов? Тестовое скрещивание по двум признакам (например, родитель мухи с голубым телом и нормальными крыльями и родитель мухи с черным телом и рудиментарными крыльями) может помочь идентифицировать рекомбинантные фенотипы. Потомство с фенотипами, отличными от родителей (например, муха с голубым телом, рудиментарными крыльями или муха с черным телом, с нормальными крыльями) указывает на то, что они являются рекомбинантными.
Соотношение фенотипов
Квадрат Пеннета — еще один инструмент, который можно использовать для идентификации всех возможных комбинаций аллелей в тестовом скрещивании.Он может предсказать генотипы и фенотипы потомства. Это диаграмма в сетках и буквах для представления аллелей. Заглавная буква (например, А) обозначает доминантный признак или генотип, а строчная буква обозначает рецессивный признак или генотип (например, а). На основе квадрата Пеннета можно определить соотношение фенотипов (а также соотношение генотипов).
Фенотипическое соотношение — это соотношение, которое можно предсказать на основе тестового скрещивания. Это можно узнать по фенотипам потомства, т.е.е. частота потомства, проявляющего различные признаки или комбинации признаков. Например, ожидаемое соотношение фенотипов дигибридного скрещивания AaBb x aabb (т. е. скрещивания, которое имеет дело с двумя признаками, например, цветом тела — A для синего, a для черного) и морфологией крыла — B для нормальных крыльев, b для рудиментарных крыльев) составляет 1:1:1:1, исходя из четырех возможных различных фенотипов: AaBb (синяя муха с нормальными крыльями), aaBb (черная муха с нормальными крыльями). ), Aabb (синяя муха с рудиментарными крыльями) и aabb (черная муха с рудиментарными крыльями).
Читать: Доминирование – Учебные пособия по биологии
Каков фенотип неполного доминирования? Как насчет кодоминирования? Скажи нам что ты думаешь! Присоединяйтесь к нашему форуму: Неполное доминирование против кодоминирования.
Примеры
Как указывалось ранее, фенотип организма относится к различным признакам, которыми обладает организм. Примерами являются черта голубых глаз (для характера цвета глаз), черта коричневой кожи (для характера цвета кожи), черта длинного хвоста (для характера хвоста), черта с пятью лепестками (для характера цветка) и т. Д.Поведение также является примером фенотипа. Например, поведение и когнитивные особенности людей с умственной отсталостью являются поведенческими фенотипами. (Ref.2)


 Исследователям необходимо проанализировать лишь небольшие участки генома под названием однонуклеотидные полиморфизмы (ОНП). Во многих их них и зашифрованы особенности внешности разных людей.
Исследователям необходимо проанализировать лишь небольшие участки генома под названием однонуклеотидные полиморфизмы (ОНП). Во многих их них и зашифрованы особенности внешности разных людей.


 1 — чуваши, 2 — коми-пермяки, 3 — коми-зыряне, 4 — марийцы равнинные, 5 — марийцы горные, 6 — мордва эрзя, 7 — мордва мокша, 8 — русские, 9 — русские (казаки-некрасовцы), 10 — русские из Нижегородской области, 11 — русские из Тверской области, 12 — русские из Ярославской области, 13 — удмурты, 14 — татары Поволжья, 15 — адыгейцы, 16 — аварцы, 17 — азербайджанцы, 18 — даргинцы, 19 — кабардинцы, 20 — карачаевцы, 21 — кумыки, 22 — лезгины, 23 — осетины, 24 — рутульцы, 25 — талыши, 26 — цахуры, 27 — турки-месхетинцы, 28 — башкиры, 29 — лесные ненцы, 30 — ханты, 31 — манси, 32 — шорцы, 33 — сибирские татары, 34 — буряты, 35 — чукчи, 36 — дунгане, 37 — эвенки Дальнего Востока, 38 — эвенки Камчатки, 39 — эвенки Охотского побережья, 40 — казахи, 41 — киргизы, 42 — коряки, 43 — нанайцы, 44 — таджики, 45 — туркмены, 46 — уйгуры, 47 — узбеки, 48 — якуты Дальнего Востока. На карте голубой цвет кружков соответствует Европейской части России; зелёный — Кавказу; фиолетовый — Западной Сибири; красный — Северной и Центральной Азии.
1 — чуваши, 2 — коми-пермяки, 3 — коми-зыряне, 4 — марийцы равнинные, 5 — марийцы горные, 6 — мордва эрзя, 7 — мордва мокша, 8 — русские, 9 — русские (казаки-некрасовцы), 10 — русские из Нижегородской области, 11 — русские из Тверской области, 12 — русские из Ярославской области, 13 — удмурты, 14 — татары Поволжья, 15 — адыгейцы, 16 — аварцы, 17 — азербайджанцы, 18 — даргинцы, 19 — кабардинцы, 20 — карачаевцы, 21 — кумыки, 22 — лезгины, 23 — осетины, 24 — рутульцы, 25 — талыши, 26 — цахуры, 27 — турки-месхетинцы, 28 — башкиры, 29 — лесные ненцы, 30 — ханты, 31 — манси, 32 — шорцы, 33 — сибирские татары, 34 — буряты, 35 — чукчи, 36 — дунгане, 37 — эвенки Дальнего Востока, 38 — эвенки Камчатки, 39 — эвенки Охотского побережья, 40 — казахи, 41 — киргизы, 42 — коряки, 43 — нанайцы, 44 — таджики, 45 — туркмены, 46 — уйгуры, 47 — узбеки, 48 — якуты Дальнего Востока. На карте голубой цвет кружков соответствует Европейской части России; зелёный — Кавказу; фиолетовый — Западной Сибири; красный — Северной и Центральной Азии.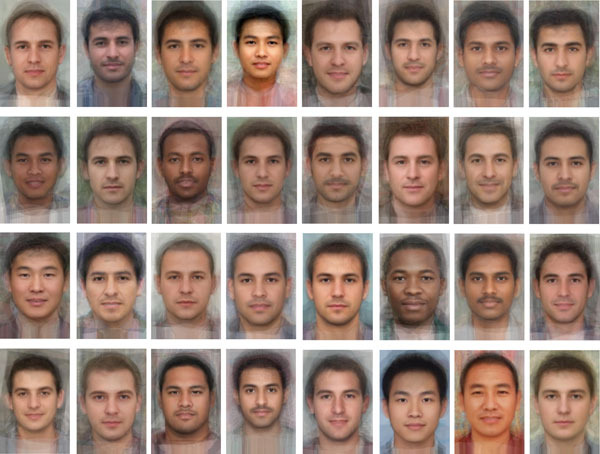
 24 SNP маркера позволяют предсказывать цвет глаз и волос, 17 SNP маркеров — цвет кожи (далее пойдёт речь только о цвете глаз и волос). Панель HIrisPlex-S была разработана на базе западноевропейских геномов (в основном геномов голландцев). И потому она с хорошей точностью работает, предсказывая цвет глаз и волос по ДНК для людей европейского происхождения. Но у людей иного происхождения генетические варианты, связанные с пигментацией волос и глаз, могут быть другими, поскольку в разных популяциях они порой возникали независимо. Проблема повышения точности европейской панели SNP маркеров особенно актуальна для России, где проживает более сотни разных народов.
24 SNP маркера позволяют предсказывать цвет глаз и волос, 17 SNP маркеров — цвет кожи (далее пойдёт речь только о цвете глаз и волос). Панель HIrisPlex-S была разработана на базе западноевропейских геномов (в основном геномов голландцев). И потому она с хорошей точностью работает, предсказывая цвет глаз и волос по ДНК для людей европейского происхождения. Но у людей иного происхождения генетические варианты, связанные с пигментацией волос и глаз, могут быть другими, поскольку в разных популяциях они порой возникали независимо. Проблема повышения точности европейской панели SNP маркеров особенно актуальна для России, где проживает более сотни разных народов.

 Итоговый список для предсказания цвета глаз включает 36 SNP, из них семь проявляют наибольшую предсказательную силу. Два SNP из этих семи были уже известны и входили в систему HIrisPlex-S, остальные пять ранее не были описаны. Четыре SNP маркера из пяти лежат в известном гене HERC2, связанном с изменчивостью цвета кожи, волос и глаз; пятый SNP расположен в межгенном регионе. Такой же анализ провели для генетических вариантов, определяющих цвет волос. Итоговый список включил 33 SNP, из которых 11 обладали наибольшей предсказательной силой. Три из них были уже известны и включены в панель HIrisPlex-S, остальные восемь ранее не описаны.
Итоговый список для предсказания цвета глаз включает 36 SNP, из них семь проявляют наибольшую предсказательную силу. Два SNP из этих семи были уже известны и входили в систему HIrisPlex-S, остальные пять ранее не были описаны. Четыре SNP маркера из пяти лежат в известном гене HERC2, связанном с изменчивостью цвета кожи, волос и глаз; пятый SNP расположен в межгенном регионе. Такой же анализ провели для генетических вариантов, определяющих цвет волос. Итоговый список включил 33 SNP, из которых 11 обладали наибольшей предсказательной силой. Три из них были уже известны и включены в панель HIrisPlex-S, остальные восемь ранее не описаны.

 Тем не менее люди (даже те, кто уверяет, что относится к геям абсолютно лояльно) подвержены этим стереотипам до такой степени, что готовы бить геев током. Статьи были опубликованы в журналах Psychological Science и The Journal of Sex Research.
Тем не менее люди (даже те, кто уверяет, что относится к геям абсолютно лояльно) подвержены этим стереотипам до такой степени, что готовы бить геев током. Статьи были опубликованы в журналах Psychological Science и The Journal of Sex Research. Результаты работы показали: при вынесении решения люди ориентировались не на фенотип человека (так как наличие или отсутствие прически на фото не влияло на «приговор» и вызывало изменение решения лишь в 4–7% случаев), а на подписи — то есть на стереотипы, — которые вызывали 24–28% вариаций, но при этом на самом деле действительность не отражали.
Результаты работы показали: при вынесении решения люди ориентировались не на фенотип человека (так как наличие или отсутствие прически на фото не влияло на «приговор» и вызывало изменение решения лишь в 4–7% случаев), а на подписи — то есть на стереотипы, — которые вызывали 24–28% вариаций, но при этом на самом деле действительность не отражали. Если мужчина при этом водит фургон и поддерживает Республиканскую партию (эксперимент, напомним, проводился в США), это окончательно снимает с него все подозрения в нетрадиционной ориентации.
Если мужчина при этом водит фургон и поддерживает Республиканскую партию (эксперимент, напомним, проводился в США), это окончательно снимает с него все подозрения в нетрадиционной ориентации. Суть игры заключалась в следующем: когда загоралась красная лампочка, испытуемый должен был нажать кнопку, в результате чего соперника било током. Соперник находился за перегородкой и был невидим, а силу тока участник мог выбирать по своему усмотрению. Целью игры было опередить соперника и нажать кнопку раньше него.
Суть игры заключалась в следующем: когда загоралась красная лампочка, испытуемый должен был нажать кнопку, в результате чего соперника било током. Соперник находился за перегородкой и был невидим, а силу тока участник мог выбирать по своему усмотрению. Целью игры было опередить соперника и нажать кнопку раньше него. Кроме того, стало очевидно, что многие из нас — даже если пытаются показаться лояльными — на самом деле относятся к гомосексуалам не так, как к традиционно ориентированным мужчинам и женщинам.
Кроме того, стало очевидно, что многие из нас — даже если пытаются показаться лояльными — на самом деле относятся к гомосексуалам не так, как к традиционно ориентированным мужчинам и женщинам. Чокмосов, С.В. Шин
Чокмосов, С.В. Шин Н. Шихвердиев, Г.Г. Хубулава, С.П. Марченко, В.В. Суворов, В.В. Зайцев, И.И. Аверкин, Р.Б. Бадуров
Н. Шихвердиев, Г.Г. Хубулава, С.П. Марченко, В.В. Суворов, В.В. Зайцев, И.И. Аверкин, Р.Б. Бадуров Главный научный сотрудник лаборатории популяционной генетики человека ФГБНУ МГНЦ, д.б.н., профессор РАН Олег Балановский выступил с докладом «2D and 3D imaging of the various populations from Russia: updating the pigmentation-predicting system» («2D и 3D фотопортреты различных популяций России: обновление системы предсказания пигментации по ДНК»).
Главный научный сотрудник лаборатории популяционной генетики человека ФГБНУ МГНЦ, д.б.н., профессор РАН Олег Балановский выступил с докладом «2D and 3D imaging of the various populations from Russia: updating the pigmentation-predicting system» («2D и 3D фотопортреты различных популяций России: обновление системы предсказания пигментации по ДНК»). Это выразилось в таких чертах, как:
Это выразилось в таких чертах, как: Таковы боги индуизма, китайские и христианские святые.
Таковы боги индуизма, китайские и христианские святые.
 Точное расположение нуклеотидов (каждый из которых состоит из фосфатной группы, сахара и основания) в гене может отличаться в разных копиях одного и того же гена. Следовательно, ген может существовать в разных формах в разных организмах. Эти различные формы известны как аллели. Точно фиксированное положение на хромосоме, которое содержит определенный ген, известно как локус.
Точное расположение нуклеотидов (каждый из которых состоит из фосфатной группы, сахара и основания) в гене может отличаться в разных копиях одного и того же гена. Следовательно, ген может существовать в разных формах в разных организмах. Эти различные формы известны как аллели. Точно фиксированное положение на хромосоме, которое содержит определенный ген, известно как локус.
 Ключевое различие между фенотипом и генотипом заключается в том, что генотип наследуется от родителей организма, а фенотип — нет.
Ключевое различие между фенотипом и генотипом заключается в том, что генотип наследуется от родителей организма, а фенотип — нет.

 Это имеет важные последствия для персонализированной медицины и подчеркивает необходимость быть осторожными, полагаясь исключительно на генотипирование.
Это имеет важные последствия для персонализированной медицины и подчеркивает необходимость быть осторожными, полагаясь исключительно на генотипирование. Этот определяемый ДНК внешний вид, или фенотип, создает семейное сходство и, по словам генетика Ричарда Спритца, «то, на что реагирует ваша бабушка, когда она говорит, что вы похожи на своего отца». Усилия генетиков по поиску фрагментов ДНК, которые определяют, как выглядит человеческое лицо — все, от формы носа до расстояния между глазами, — в последние годы активизировались, и был достигнут прогресс. Теперь ученые могут с некоторой уверенностью использовать цепочку ДНК для определения вероятного цвета волос и глаз человека, а также пигментации кожи и происхождения.Генетик из Пенсильванского государственного университета Марк Шрайвер предпринял то, что он описывает как «первую попытку создания лицевых композитов из ДНК» с «предварительными, но, безусловно, многообещающими результатами».
Этот определяемый ДНК внешний вид, или фенотип, создает семейное сходство и, по словам генетика Ричарда Спритца, «то, на что реагирует ваша бабушка, когда она говорит, что вы похожи на своего отца». Усилия генетиков по поиску фрагментов ДНК, которые определяют, как выглядит человеческое лицо — все, от формы носа до расстояния между глазами, — в последние годы активизировались, и был достигнут прогресс. Теперь ученые могут с некоторой уверенностью использовать цепочку ДНК для определения вероятного цвета волос и глаз человека, а также пигментации кожи и происхождения.Генетик из Пенсильванского государственного университета Марк Шрайвер предпринял то, что он описывает как «первую попытку создания лицевых композитов из ДНК» с «предварительными, но, безусловно, многообещающими результатами». Физический портрет подозреваемого может быть разработан на основе ДНК, оставленной на месте происшествия, когда не было свидетелей преступления.После пожаров или других катастроф ДНК неопознанных тел может сделать их узнаваемыми для членов семьи. Кроме того, ДНК из фрагмента кости может помочь визуализировать и идентифицировать людей в массовых захоронениях. Хотя большинство генетиков подчеркивают ограниченность исследований фенотипов, полиция и судебные следователи уже могут обратиться к горстке частных компаний, которые утверждают, что могут использовать ДНК для точного предсказания внешности человека. Итак, каково состояние исследований фенотипа? Может ли судебно-медицинский следователь реально рассчитывать получить точное изображение человека из фрагмента ДНК?
Физический портрет подозреваемого может быть разработан на основе ДНК, оставленной на месте происшествия, когда не было свидетелей преступления.После пожаров или других катастроф ДНК неопознанных тел может сделать их узнаваемыми для членов семьи. Кроме того, ДНК из фрагмента кости может помочь визуализировать и идентифицировать людей в массовых захоронениях. Хотя большинство генетиков подчеркивают ограниченность исследований фенотипов, полиция и судебные следователи уже могут обратиться к горстке частных компаний, которые утверждают, что могут использовать ДНК для точного предсказания внешности человека. Итак, каково состояние исследований фенотипа? Может ли судебно-медицинский следователь реально рассчитывать получить точное изображение человека из фрагмента ДНК? Хотя генетики с осторожностью относятся к преувеличению прогресса в создании точного физического образа человека на основе ДНК, существует общее мнение, что понимание основ фенотипов значительно продвинулось вперед за последнее десятилетие. Теперь ученые могут использовать ДНК для определения с вероятностью более 75 процентов происхождения человека, цвета глаз и волос.
Хотя генетики с осторожностью относятся к преувеличению прогресса в создании точного физического образа человека на основе ДНК, существует общее мнение, что понимание основ фенотипов значительно продвинулось вперед за последнее десятилетие. Теперь ученые могут использовать ДНК для определения с вероятностью более 75 процентов происхождения человека, цвета глаз и волос. Кидд расширяет панель AISNP, чтобы включить больше географических регионов. Определение происхождения человека — это часть генетической головоломки, которая определяет, как выглядит этот человек, и это важная часть более широких усилий по точному изображению конкретного лица по ДНК. Несмотря на прогресс, которого он добивается, Кидд сказал, что ему еще предстоит пройти долгий путь. «С такими исследованиями, которыми я занимаюсь, — сказал он, — я никогда не закончу».
Кидд расширяет панель AISNP, чтобы включить больше географических регионов. Определение происхождения человека — это часть генетической головоломки, которая определяет, как выглядит этот человек, и это важная часть более широких усилий по точному изображению конкретного лица по ДНК. Несмотря на прогресс, которого он добивается, Кидд сказал, что ему еще предстоит пройти долгий путь. «С такими исследованиями, которыми я занимаюсь, — сказал он, — я никогда не закончу». Нашей целью сейчас является определение реального цвета, как значение RGB в Adobe Photoshop». Что вдохновило ее на определение реального цвета по ДНК, так это просьба молекулярного генетика Тури Кинга определить цвет глаз и волос Ричарда III, останки которого были найдены под парковкой в Лестере, Англия, в 2012 году.Кинг, который использовал митохондриальную ДНК, чтобы подтвердить, что останки принадлежат Ричарду III, обратился к Уолшу, чтобы определить, какой из портретов короля — все они были написаны после того, как он был убит в бою в 1485 году — был наиболее точным. Основываясь на анализе фенотипа Уолша, Кинг определил, что одна из самых ранних картин Ричарда III, «Портрет в арочной раме» 1510 года, лучше всего соответствует генетической информации.
Нашей целью сейчас является определение реального цвета, как значение RGB в Adobe Photoshop». Что вдохновило ее на определение реального цвета по ДНК, так это просьба молекулярного генетика Тури Кинга определить цвет глаз и волос Ричарда III, останки которого были найдены под парковкой в Лестере, Англия, в 2012 году.Кинг, который использовал митохондриальную ДНК, чтобы подтвердить, что останки принадлежат Ричарду III, обратился к Уолшу, чтобы определить, какой из портретов короля — все они были написаны после того, как он был убит в бою в 1485 году — был наиболее точным. Основываясь на анализе фенотипа Уолша, Кинг определил, что одна из самых ранних картин Ричарда III, «Портрет в арочной раме» 1510 года, лучше всего соответствует генетической информации. Потому что я всегда мог сказать кому-нибудь «голубой» или «блондин», а они бы сказали: «Мне нужно увидеть это физически». Вот над чем я сейчас работаю. Я хочу добиться такого результата». Уолш собрал данные о фенотипах ДНК от 2000 человек из Ирландии, Греции и США и в настоящее время собирает данные еще от 3000 человек из тех же стран, чтобы создать базу данных фенотип-генотип и модель прогнозирования.Для судебно-медицинских целей она хотела бы иметь возможность начать с «пустого человека» и по образцу ДНК определить настоящую пигментацию глаз, волос и кожи.
Потому что я всегда мог сказать кому-нибудь «голубой» или «блондин», а они бы сказали: «Мне нужно увидеть это физически». Вот над чем я сейчас работаю. Я хочу добиться такого результата». Уолш собрал данные о фенотипах ДНК от 2000 человек из Ирландии, Греции и США и в настоящее время собирает данные еще от 3000 человек из тех же стран, чтобы создать базу данных фенотип-генотип и модель прогнозирования.Для судебно-медицинских целей она хотела бы иметь возможность начать с «пустого человека» и по образцу ДНК определить настоящую пигментацию глаз, волос и кожи. «Предыдущая работа, связанная с линейным измерением между ориентирами, не сработала. Итак, его новый подход состоит в том, чтобы сосредоточиться на ориентирах, которые имеют самый высокий генетический компонент наследуемости. «Вот с чего мы начнем», — сказал Спритц. «Мы очень сильно скрестили пальцы, чтобы это выглядело лучше».
«Предыдущая работа, связанная с линейным измерением между ориентирами, не сработала. Итак, его новый подход состоит в том, чтобы сосредоточиться на ориентирах, которые имеют самый высокий генетический компонент наследуемости. «Вот с чего мы начнем», — сказал Спритц. «Мы очень сильно скрестили пальцы, чтобы это выглядело лучше». «Было бы ошибкой утверждать, что это слишком сложный процесс для нашего понимания. Тело это понимает. Есть все эти сложные взаимодействия, и они считываются телом. Есть ли в этом что-то особенное, что означает, что мы не сможем это понять?»
«Было бы ошибкой утверждать, что это слишком сложный процесс для нашего понимания. Тело это понимает. Есть все эти сложные взаимодействия, и они считываются телом. Есть ли в этом что-то особенное, что означает, что мы не сможем это понять?» «Есть некоторые генетики, которые относительно просты, например, болезнь», — сказал Спритц. «Есть промежуточные значения, такие как рост человека, а есть невообразимо сложные, такие как определение формы и черт лица».
«Есть некоторые генетики, которые относительно просты, например, болезнь», — сказал Спритц. «Есть промежуточные значения, такие как рост человека, а есть невообразимо сложные, такие как определение формы и черт лица».
 Примечательно, и там, где криминалистическое фенотипирование ДНК показывает большие перспективы, это относится к (гораздо) меньшей группе потенциальных подозреваемых, которые соответствуют характеристикам внешности, предсказанным ДНК по пятну с места преступления или по останкам умершего человека.Ожидается, что при наличии достаточного финансирования будущие исследования, направленные на лучшее понимание генетической основы внешности человека, приведут к значительно более подробному описанию внешности неизвестного человека на основе ДНК, что повысит ценность полицейских расследований уголовных дел и дел о пропавших без вести с участием неизвестных.
Примечательно, и там, где криминалистическое фенотипирование ДНК показывает большие перспективы, это относится к (гораздо) меньшей группе потенциальных подозреваемых, которые соответствуют характеристикам внешности, предсказанным ДНК по пятну с места преступления или по останкам умершего человека.Ожидается, что при наличии достаточного финансирования будущие исследования, направленные на лучшее понимание генетической основы внешности человека, приведут к значительно более подробному описанию внешности неизвестного человека на основе ДНК, что повысит ценность полицейских расследований уголовных дел и дел о пропавших без вести с участием неизвестных.
 И это то, что я также говорю студентам: «Не прыгайте сразу в аспирантуру, это от 4 до 5 лет вашей жизни, так что вы должны сдерживаться и действительно, действительно убедиться, что тема, которую вы повторный выбор — это то, что вы хотите сделать. Я буквально только что погуглил «ДНК судебной экспертизы внешности» и наткнулся на Dr.Манфред Кайзер. Я отправил ему электронное письмо, а остальное уже история».
И это то, что я также говорю студентам: «Не прыгайте сразу в аспирантуру, это от 4 до 5 лет вашей жизни, так что вы должны сдерживаться и действительно, действительно убедиться, что тема, которую вы повторный выбор — это то, что вы хотите сделать. Я буквально только что погуглил «ДНК судебной экспертизы внешности» и наткнулся на Dr.Манфред Кайзер. Я отправил ему электронное письмо, а остальное уже история». Эта информация может быть использована в контексте судебно-медицинской экспертизы для предоставления правоохранительным органам ключевой разведывательной информации, такой как цвет глаз, волос или кожи. Хотя эта информация не является исчерпывающей, она может быть использована для того, чтобы дать указания правоохранительным органам в качестве следственного ориентира в сложных делах о пропавших без вести или в ситуации массового бедствия.
Эта информация может быть использована в контексте судебно-медицинской экспертизы для предоставления правоохранительным органам ключевой разведывательной информации, такой как цвет глаз, волос или кожи. Хотя эта информация не является исчерпывающей, она может быть использована для того, чтобы дать указания правоохранительным органам в качестве следственного ориентира в сложных делах о пропавших без вести или в ситуации массового бедствия.
 У человека каталог OMIM ® (Online Mendelian Inheritance in Man, http://omim.org/), в котором собраны генетические детерминанты фенотипов, связанных с заболеванием, насчитывает более 4300 записей и в общей сложности 2493 опубликованных полногеномных ассоциативных исследования. (GWAS) обнаружили множество участков в геноме, которые статистически связаны со сложными признаками (Welter et al., 2014). Поскольку обнаружение причинно-следственных связей между генетическими и фенотипическими вариациями ускоряется, пересмотр наших концептуальных инструментов может помочь нам найти объединяющие принципы в рое данных. Здесь мы размышляем о взаимосвязи между генотипами и фенотипами и адресуем это эссе биологам, которые готовы попытаться бросить вызов своему нынешнему пониманию фенотипов. Выделим одну полезную точку зрения — дифференциальную. Затем мы показываем, что эта простая структура остается полезной в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды.
У человека каталог OMIM ® (Online Mendelian Inheritance in Man, http://omim.org/), в котором собраны генетические детерминанты фенотипов, связанных с заболеванием, насчитывает более 4300 записей и в общей сложности 2493 опубликованных полногеномных ассоциативных исследования. (GWAS) обнаружили множество участков в геноме, которые статистически связаны со сложными признаками (Welter et al., 2014). Поскольку обнаружение причинно-следственных связей между генетическими и фенотипическими вариациями ускоряется, пересмотр наших концептуальных инструментов может помочь нам найти объединяющие принципы в рое данных. Здесь мы размышляем о взаимосвязи между генотипами и фенотипами и адресуем это эссе биологам, которые готовы попытаться бросить вызов своему нынешнему пониманию фенотипов. Выделим одну полезную точку зрения — дифференциальную. Затем мы показываем, что эта простая структура остается полезной в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды.
 На самом деле ген сам по себе не может ни вызвать наблюдаемый фенотипический признак, ни быть необходимым и достаточным для появления наблюдаемых признаков.Генам нужна клеточная среда, совместное действие множества других генов, а также определенные физико-химические условия, чтобы оказывать заметное влияние на организмы (рис. 1В). Например, пигментация каштановых волос у одного человека является результатом не только действия генов, кодирующих ферменты, синтезирующие пигмент, но и наличия клеток, продуцирующих пигменты соответствующих молекул субстрата (таких как тирозин для меланина), и количества полученного вещества. солнечный свет (Liu et al., 2013). Таким образом, генетический редукционистский подход, который исследует лишь несколько генетических параметров среди множества причинных факторов, бесполезен для полного решения широкого вопроса о том, что делает волосы коричневыми, что порождает определенную биологическую структуру или процесс в целом.Тем не менее, генетический редукционизм может быть совершенно подходящим для идентификации генетических локусов, где изменение вызывает фенотипические различия (рис.
На самом деле ген сам по себе не может ни вызвать наблюдаемый фенотипический признак, ни быть необходимым и достаточным для появления наблюдаемых признаков.Генам нужна клеточная среда, совместное действие множества других генов, а также определенные физико-химические условия, чтобы оказывать заметное влияние на организмы (рис. 1В). Например, пигментация каштановых волос у одного человека является результатом не только действия генов, кодирующих ферменты, синтезирующие пигмент, но и наличия клеток, продуцирующих пигменты соответствующих молекул субстрата (таких как тирозин для меланина), и количества полученного вещества. солнечный свет (Liu et al., 2013). Таким образом, генетический редукционистский подход, который исследует лишь несколько генетических параметров среди множества причинных факторов, бесполезен для полного решения широкого вопроса о том, что делает волосы коричневыми, что порождает определенную биологическую структуру или процесс в целом.Тем не менее, генетический редукционизм может быть совершенно подходящим для идентификации генетических локусов, где изменение вызывает фенотипические различия (рис. 1С). Различие в цвете волос между двумя людьми в некоторых случаях может быть связано с их генетическими различиями. Заметим, однако, что не все фенотипические изменения можно отнести к генетическим изменениям. Различия в цвете волос также могут быть вызваны негенетическими факторами, такими как возраст, интенсивность солнечного излучения или окраска волос, или сочетанием как генетических, так и негенетических различий.
1С). Различие в цвете волос между двумя людьми в некоторых случаях может быть связано с их генетическими различиями. Заметим, однако, что не все фенотипические изменения можно отнести к генетическим изменениям. Различия в цвете волос также могут быть вызваны негенетическими факторами, такими как возраст, интенсивность солнечного излучения или окраска волос, или сочетанием как генетических, так и негенетических различий. Долгое время некоторые генетики могли думать, что они анализируют морфогенетические механизмы, лежащие в основе формирования фенотипических признаков, в то время как их экспериментальный подход фактически выявлял гены, отсутствие или изменение которых (мутации, делеции, дупликации, перестройки и т. д.) приводит к фенотипические различия (сравните рисунок 1А с рисунком 1С). На самом деле предложение «ваши волосы каштановые» можно интерпретировать либо как абсолютное наблюдение (описание определенного набора молекул, содержащих определенные уровни темного пигмента эумеланина и светлого пигмента феомеланина), либо как имплицитную ссылку на другие возможности. (он коричневый, а не другого цвета).Заблуждения возникают из-за того, что фенотипы обычно определяются относительно возможностей, которые не сформулированы явно. Наше сознание и наш язык часто склонны путать объекты, изменчивость которых рассматривается, с самой изменчивостью (Keller, 2010), и важно напомнить, что в генетике интересующие нас объекты (например, данный генотип, аллель или фенотип) заслуживают определения 90 318 относительно 90 319 по отношению к другому эталонному состоянию.
Долгое время некоторые генетики могли думать, что они анализируют морфогенетические механизмы, лежащие в основе формирования фенотипических признаков, в то время как их экспериментальный подход фактически выявлял гены, отсутствие или изменение которых (мутации, делеции, дупликации, перестройки и т. д.) приводит к фенотипические различия (сравните рисунок 1А с рисунком 1С). На самом деле предложение «ваши волосы каштановые» можно интерпретировать либо как абсолютное наблюдение (описание определенного набора молекул, содержащих определенные уровни темного пигмента эумеланина и светлого пигмента феомеланина), либо как имплицитную ссылку на другие возможности. (он коричневый, а не другого цвета).Заблуждения возникают из-за того, что фенотипы обычно определяются относительно возможностей, которые не сформулированы явно. Наше сознание и наш язык часто склонны путать объекты, изменчивость которых рассматривается, с самой изменчивостью (Keller, 2010), и важно напомнить, что в генетике интересующие нас объекты (например, данный генотип, аллель или фенотип) заслуживают определения 90 318 относительно 90 319 по отношению к другому эталонному состоянию.
 Мы покажем, что отношения ГП — это гораздо больше, чем простое и слабо определенное взаимодействие между двумя уровнями организации: это причинно-следственная связь, которая облегчает наше понимание фенотипического разнообразия.
Мы покажем, что отношения ГП — это гораздо больше, чем простое и слабо определенное взаимодействие между двумя уровнями организации: это причинно-следственная связь, которая облегчает наше понимание фенотипического разнообразия. Здесь акцент делается не на сам ген, как он определен в геномных базах данных, а скорее на индивидуальном функциональном разделении генома на определяющие различия локусы. Генотипическая часть родства GP может принимать форму различных аллелей: разные кодоны, кодирующие разные аминокислоты, вставки/делеции в последовательности, кодирующей белок, расходящиеся версии конкретного регуляторного элемента cis-, наличие/отсутствие вставок транспозонов. , количество копий генов в кластере генов, склонных к структурной изменчивости, и т. д.В геноме не все нуклеотидные участки связаны с фенотипической изменчивостью. Например, вероятно, существуют фрагменты нуклеотидных последовательностей, в том числе так называемая мусорная ДНК (Graur et al., 2015), присутствие которых не оказывает никакого влияния на наблюдаемые характеристики организма, кроме того, что они реплицируются и, возможно, транскрибируются. Существуют также генетические локусы, которые могли быть связаны с фенотипической изменчивостью в прошлом и больше не связаны с фенотипической изменчивостью.
Здесь акцент делается не на сам ген, как он определен в геномных базах данных, а скорее на индивидуальном функциональном разделении генома на определяющие различия локусы. Генотипическая часть родства GP может принимать форму различных аллелей: разные кодоны, кодирующие разные аминокислоты, вставки/делеции в последовательности, кодирующей белок, расходящиеся версии конкретного регуляторного элемента cis-, наличие/отсутствие вставок транспозонов. , количество копий генов в кластере генов, склонных к структурной изменчивости, и т. д.В геноме не все нуклеотидные участки связаны с фенотипической изменчивостью. Например, вероятно, существуют фрагменты нуклеотидных последовательностей, в том числе так называемая мусорная ДНК (Graur et al., 2015), присутствие которых не оказывает никакого влияния на наблюдаемые характеристики организма, кроме того, что они реплицируются и, возможно, транскрибируются. Существуют также генетические локусы, которые могли быть связаны с фенотипической изменчивостью в прошлом и больше не связаны с фенотипической изменчивостью. Например, генетическая изменчивость в кодирующих областях, связывающих гистоновую ДНК, могла быть важной во время ранней эволюции эукариотических клеток, но эти генетические локусы больше не содержат фенотипически релевантных вариаций, кроме летальных мутаций. Таким образом, в геноме есть участки нуклеотидов, которые абсолютно необходимы для жизни, но сами по себе не содержат жизнеспособных фенотипически релевантных вариаций.
Например, генетическая изменчивость в кодирующих областях, связывающих гистоновую ДНК, могла быть важной во время ранней эволюции эукариотических клеток, но эти генетические локусы больше не содержат фенотипически релевантных вариаций, кроме летальных мутаций. Таким образом, в геноме есть участки нуклеотидов, которые абсолютно необходимы для жизни, но сами по себе не содержат жизнеспособных фенотипически релевантных вариаций. , 1931). В других случаях причинное генетическое изменение связано с симбионтными бактериями: температурная толерантность тли может различаться у разных особей из-за точечной мутации в их бактериальном симбионте (Dunbar et al., 2007). Определенные фенотипические эффекты могут проявляться и на более высоком уровне, чем организм, несущий генетическое изменение (Dawkins, 1982), например, социальная организация муравьиной колонии (Wang et al., 2013).
, 1931). В других случаях причинное генетическое изменение связано с симбионтными бактериями: температурная толерантность тли может различаться у разных особей из-за точечной мутации в их бактериальном симбионте (Dunbar et al., 2007). Определенные фенотипические эффекты могут проявляться и на более высоком уровне, чем организм, несущий генетическое изменение (Dawkins, 1982), например, социальная организация муравьиной колонии (Wang et al., 2013). Следует отметить, что изменчивость фенотипа не всегда сразу следует за появлением новой вызывающей мутации, но может появиться позже из-за единичного набора аллелей, сегрегирующих в популяции.Например, новый фенотип редуцированных панцирных пластин появился в популяции пресноводной колюшки, когда рецессивный аллель EDA , уже присутствующий на криптических уровнях, оказался в гомозиготном состоянии у одной особи (Colosimo et al., 2005; Jones et al., 2012). Главный концептуальный прорыв, сделанный Чарльзом Дарвином, заключался в том, чтобы связать изменчивость среди особей внутри скрещивающейся группы (различие № 2) с изменчивостью между таксономическими группами в пространстве и времени (различие № 1; Левонтин, 1974а).
Следует отметить, что изменчивость фенотипа не всегда сразу следует за появлением новой вызывающей мутации, но может появиться позже из-за единичного набора аллелей, сегрегирующих в популяции.Например, новый фенотип редуцированных панцирных пластин появился в популяции пресноводной колюшки, когда рецессивный аллель EDA , уже присутствующий на криптических уровнях, оказался в гомозиготном состоянии у одной особи (Colosimo et al., 2005; Jones et al., 2012). Главный концептуальный прорыв, сделанный Чарльзом Дарвином, заключался в том, чтобы связать изменчивость среди особей внутри скрещивающейся группы (различие № 2) с изменчивостью между таксономическими группами в пространстве и времени (различие № 1; Левонтин, 1974а). Тем не менее, у этих женщин риск развития рака груди или яичников к 70 годам составляет до 80% по сравнению с женщинами, имеющими две копии дикого типа BRCA1 , из-за появления дополнительных вредных мутаций в диком типе BRCA1 . аллель в их соматических клетках молочной железы (Narod and Foulkes, 2004).
Тем не менее, у этих женщин риск развития рака груди или яичников к 70 годам составляет до 80% по сравнению с женщинами, имеющими две копии дикого типа BRCA1 , из-за появления дополнительных вредных мутаций в диком типе BRCA1 . аллель в их соматических клетках молочной железы (Narod and Foulkes, 2004). Этот пример показывает, что причинно-следственная связь между генетическим изменением и связанным с ним фенотипическим изменением может скрывать множество встроенных параметров (таких как медицинская практика в случае фенилаланингидроксилазы) в рамках предположения при прочих равных «при прочих равных условиях.
Этот пример показывает, что причинно-следственная связь между генетическим изменением и связанным с ним фенотипическим изменением может скрывать множество встроенных параметров (таких как медицинская практика в случае фенилаланингидроксилазы) в рамках предположения при прочих равных «при прочих равных условиях. 2А).Он указал средний генотип популяции как точку в пространстве всех возможных генотипов (пространство G) и средний фенотип той же популяции как соответствующую точку в пространстве всех возможных фенотипов (пространство P). Таким образом, эволюционный процесс был разбит на четыре этапа: (1) средний фенотип возникает в результате развития различных генотипов в различных условиях; (2) миграция, спаривание и естественный отбор действуют в пространстве P, чтобы изменить средний фенотип исходной популяции на средний фенотип особей, которые будут иметь потомство; (3) идентичность успешных родителей определяет, какие генотипы сохраняются; и (4) генетические процессы, такие как мутация и рекомбинация, изменяют положение в G-пространстве.
2А).Он указал средний генотип популяции как точку в пространстве всех возможных генотипов (пространство G) и средний фенотип той же популяции как соответствующую точку в пространстве всех возможных фенотипов (пространство P). Таким образом, эволюционный процесс был разбит на четыре этапа: (1) средний фенотип возникает в результате развития различных генотипов в различных условиях; (2) миграция, спаривание и естественный отбор действуют в пространстве P, чтобы изменить средний фенотип исходной популяции на средний фенотип особей, которые будут иметь потомство; (3) идентичность успешных родителей определяет, какие генотипы сохраняются; и (4) генетические процессы, такие как мутация и рекомбинация, изменяют положение в G-пространстве. В третьем представлении, предложенном Вагнером (1996; рис. 2С), отдельные гены связаны с отдельными признаками.
В третьем представлении, предложенном Вагнером (1996; рис. 2С), отдельные гены связаны с отдельными признаками. Из-за двух запутанных определений гена, либо как кодирующего белок, либо как вызывающего фенотипическое изменение (Griffiths and Stotz, 2013), легко перейти от дифференциального взгляда к недифференциальному взгляду на отношения GP.
Из-за двух запутанных определений гена, либо как кодирующего белок, либо как вызывающего фенотипическое изменение (Griffiths and Stotz, 2013), легко перейти от дифференциального взгляда к недифференциальному взгляду на отношения GP. Рихард Левонтин и Гюнтер Вагнер определили характеры как элементы внутри организма, которые отвечают на адаптивные вызовы и представляют собой квазинезависимые единицы эволюционных изменений (Левонтин, 1978; Вагнер, 2000).Их определение касается абсолютных признаков, наблюдаемых у отдельных организмов (например, форма крыла или количество пальцев у особи), и поэтому далеко от дифференциального взгляда. Здесь, чтобы лучше понять эволюцию и фенотипическое разнообразие живого мира, мы предлагаем разложить наблюдаемые признаки организма на множество элементарных вариаций ГП, которые накапливались в течение многих поколений, начиная с исходного состояния. Мы настаиваем на том, что с этой точки зрения персонажи являются не конкретными объектами (например, кожей), а абстрактными объектами, определяемыми существованием различий между двумя возможными наблюдаемыми состояниями (например, цветом кожи).В качестве аналогии можно представить два способа изготовления изношенной кожаной обуви определенной формы.
Рихард Левонтин и Гюнтер Вагнер определили характеры как элементы внутри организма, которые отвечают на адаптивные вызовы и представляют собой квазинезависимые единицы эволюционных изменений (Левонтин, 1978; Вагнер, 2000).Их определение касается абсолютных признаков, наблюдаемых у отдельных организмов (например, форма крыла или количество пальцев у особи), и поэтому далеко от дифференциального взгляда. Здесь, чтобы лучше понять эволюцию и фенотипическое разнообразие живого мира, мы предлагаем разложить наблюдаемые признаки организма на множество элементарных вариаций ГП, которые накапливались в течение многих поколений, начиная с исходного состояния. Мы настаиваем на том, что с этой точки зрения персонажи являются не конкретными объектами (например, кожей), а абстрактными объектами, определяемыми существованием различий между двумя возможными наблюдаемыми состояниями (например, цветом кожи).В качестве аналогии можно представить два способа изготовления изношенной кожаной обуви определенной формы. Можно либо собрать разные атомы в одну организацию, либо можно купить в магазине ботинок, а затем подвергнуть его ряду механических воздействий. Мы естественно склонны сравнивать организмы с машинами и мыслить в терминах частей, которые должны быть собраны, чтобы создать функциональное целое. Однако безудержной метафоры дизайнера или создателя недостаточно для понимания происхождения современных организмов (Coen, 2012).Чтобы понять фенотипические особенности данного организма, более эффективно разложить его на абстрактные изменения, которые происходили последовательно на протяжении эволюционного времени, а не на протяжении времени развития. Исходное состояние является гипотетическим предком изучаемого организма.
Можно либо собрать разные атомы в одну организацию, либо можно купить в магазине ботинок, а затем подвергнуть его ряду механических воздействий. Мы естественно склонны сравнивать организмы с машинами и мыслить в терминах частей, которые должны быть собраны, чтобы создать функциональное целое. Однако безудержной метафоры дизайнера или создателя недостаточно для понимания происхождения современных организмов (Coen, 2012).Чтобы понять фенотипические особенности данного организма, более эффективно разложить его на абстрактные изменения, которые происходили последовательно на протяжении эволюционного времени, а не на протяжении времени развития. Исходное состояние является гипотетическим предком изучаемого организма. ), связанные с генетической разницей. Например, мутация V370A рецептора EDAR связана не только с толщиной волос, но и с изменением плотности потовых и молочных желез в азиатских популяциях (Камберов и др., 2013). Отношения GP в таких случаях являются один-ко-многим. Рассмотрение кожи и глаза как независимых анатомических модулей человеческого тела может показаться уместным для многих эволюционных изменений, но несколько неадекватным в тех случаях, когда эти два органа развили новый признак пигментации одновременно посредством единственной мутации в гене SLC45A2 (Liu и другие., 2013). Рассуждения в терминах взаимосвязей ГП снимают проблему нахождения соответствующей декомпозиции на элементарные анатомические структуры. Сами элементарные отношения ГП представляются адекватными полунезависимыми модулями, комбинация которых может объяснить наблюдаемые характеристики организма.
), связанные с генетической разницей. Например, мутация V370A рецептора EDAR связана не только с толщиной волос, но и с изменением плотности потовых и молочных желез в азиатских популяциях (Камберов и др., 2013). Отношения GP в таких случаях являются один-ко-многим. Рассмотрение кожи и глаза как независимых анатомических модулей человеческого тела может показаться уместным для многих эволюционных изменений, но несколько неадекватным в тех случаях, когда эти два органа развили новый признак пигментации одновременно посредством единственной мутации в гене SLC45A2 (Liu и другие., 2013). Рассуждения в терминах взаимосвязей ГП снимают проблему нахождения соответствующей декомпозиции на элементарные анатомические структуры. Сами элементарные отношения ГП представляются адекватными полунезависимыми модулями, комбинация которых может объяснить наблюдаемые характеристики организма. Доступны экспериментальные подходы для разложения данного фенотипического различия на соответствующие более тонкие подвариации. Например, скрещивание растений с разной формой листьев дает потомство, которое демонстрирует составной набор промежуточных форм листьев. Анализ основных компонентов выявил элементарные изменения формы листа, которые вместе могут объяснить разницу в форме между родительскими линиями и, по-видимому, вызваны различными геномными областями (Langlade et al., 2005). Это в какой-то степени предполагает, что «сумма затемняет части.«То, что мы традиционно считаем сложными чертами, можно составить из более простых черт, более поддающихся генетическому анализу. Другим ярким примером является пигментация брюшка в группе Drosophila dunni . Взятые как одна переменная, уровни пигментации демонстрируют сложную генетическую архитектуру, но разложение взрослых паттернов на анатомические субъединицы раскрывает дискретный генетический контроль для каждого субпризнака (Hollocher et al.
Доступны экспериментальные подходы для разложения данного фенотипического различия на соответствующие более тонкие подвариации. Например, скрещивание растений с разной формой листьев дает потомство, которое демонстрирует составной набор промежуточных форм листьев. Анализ основных компонентов выявил элементарные изменения формы листа, которые вместе могут объяснить разницу в форме между родительскими линиями и, по-видимому, вызваны различными геномными областями (Langlade et al., 2005). Это в какой-то степени предполагает, что «сумма затемняет части.«То, что мы традиционно считаем сложными чертами, можно составить из более простых черт, более поддающихся генетическому анализу. Другим ярким примером является пигментация брюшка в группе Drosophila dunni . Взятые как одна переменная, уровни пигментации демонстрируют сложную генетическую архитектуру, но разложение взрослых паттернов на анатомические субъединицы раскрывает дискретный генетический контроль для каждого субпризнака (Hollocher et al. Кодирующие экзоны представлены темными прямоугольниками, а нетранслируемые экзоны — белыми прямоугольниками.Адаптировано из Linnen et al. (2013).
Кодирующие экзоны представлены темными прямоугольниками, а нетранслируемые экзоны — белыми прямоугольниками.Адаптировано из Linnen et al. (2013).
 , 2007; Рисунок 4Б). Эффект мутации MC1R виден только в присутствии ассоциированного со светлым цветом производного гаплотипа Agouti . Здесь считается, что локус Mc1R эпистатически взаимодействует с локусом Agouti . В этом случае мы предполагаем, что отношение GP включает не одно фенотипическое различие, а два 90 318 возможных 90 319 фенотипических различий (изменение пигментации шерсти или полное отсутствие изменений). Выбор между этими двумя фенотипическими различиями определяется генетическим фоном (здесь локус Agouti ).Дифференциальный взгляд, таким образом, остается относительно простым для взаимодействия двух локусов: зависимость фенотипа от контекста транслируется в выбор между двумя возможными фенотипическими различиями. Мы предполагаем, что отношения GP, включающие мутацию, подверженную множественным эпистатическим взаимодействиям, должны включать все возможные фенотипические различия, которые могут возникнуть в результате мутации во всех генетических фонах.
, 2007; Рисунок 4Б). Эффект мутации MC1R виден только в присутствии ассоциированного со светлым цветом производного гаплотипа Agouti . Здесь считается, что локус Mc1R эпистатически взаимодействует с локусом Agouti . В этом случае мы предполагаем, что отношение GP включает не одно фенотипическое различие, а два 90 318 возможных 90 319 фенотипических различий (изменение пигментации шерсти или полное отсутствие изменений). Выбор между этими двумя фенотипическими различиями определяется генетическим фоном (здесь локус Agouti ).Дифференциальный взгляд, таким образом, остается относительно простым для взаимодействия двух локусов: зависимость фенотипа от контекста транслируется в выбор между двумя возможными фенотипическими различиями. Мы предполагаем, что отношения GP, включающие мутацию, подверженную множественным эпистатическим взаимодействиям, должны включать все возможные фенотипические различия, которые могут возникнуть в результате мутации во всех генетических фонах. роста по сравнению с диким типом в нормальных почвах (Gujas et al., 2012). Взаимодействия GxE обычно анализируют в форме нормы реакции , которая представляет все наблюдаемые черты одного генотипа в различных средах (Johannsen, 1911; Sarkar, 1999).В случае взаимодействий GxE мы предполагаем, что отношения GP должны включать все возможные фенотипические изменения , которые могут быть вызваны ассоциированными генетическими изменениями в различных экспериментальных условиях. Таким образом, связанное с этим фенотипическое изменение представляет собой разницу между двумя нормами реакции. Примером из учебника является изменение правила температуры и размера у C. elegans . Как и большинство других животных, нематоды C. elegans увеличиваются в размерах при низкой температуре, но лабораторный штамм дикого типа C.elegans , происходящий с Гавайских островов, не показывает изменений в размерах тела при различных температурах. Аминокислотное изменение в кальций-связывающем белке ответственно за снижение способности гавайского штамма увеличиваться в размерах при низкой температуре (Kammenga et al.
роста по сравнению с диким типом в нормальных почвах (Gujas et al., 2012). Взаимодействия GxE обычно анализируют в форме нормы реакции , которая представляет все наблюдаемые черты одного генотипа в различных средах (Johannsen, 1911; Sarkar, 1999).В случае взаимодействий GxE мы предполагаем, что отношения GP должны включать все возможные фенотипические изменения , которые могут быть вызваны ассоциированными генетическими изменениями в различных экспериментальных условиях. Таким образом, связанное с этим фенотипическое изменение представляет собой разницу между двумя нормами реакции. Примером из учебника является изменение правила температуры и размера у C. elegans . Как и большинство других животных, нематоды C. elegans увеличиваются в размерах при низкой температуре, но лабораторный штамм дикого типа C.elegans , происходящий с Гавайских островов, не показывает изменений в размерах тела при различных температурах. Аминокислотное изменение в кальций-связывающем белке ответственно за снижение способности гавайского штамма увеличиваться в размерах при низкой температуре (Kammenga et al. , 2007). Здесь норма реакции (представляющая размер тела нематоды в диапазоне температур) различается между нематодами, и связанное с ней отношение GP охватывает разницу между этими двумя наклонами.
, 2007). Здесь норма реакции (представляющая размер тела нематоды в диапазоне температур) различается между нематодами, и связанное с ней отношение GP охватывает разницу между этими двумя наклонами. Для того чтобы дифференцированный взгляд был приемлемым, мы советуем не рассматривать все возможные генетические предпосылки и условия окружающей среды, а ограничивать возможности потенциальной средой и выделять аллели, которые имеют отношение к интересующей популяции (Sober, 1988).
Для того чтобы дифференцированный взгляд был приемлемым, мы советуем не рассматривать все возможные генетические предпосылки и условия окружающей среды, а ограничивать возможности потенциальной средой и выделять аллели, которые имеют отношение к интересующей популяции (Sober, 1988). Однако мы выделяем некоторые случаи, когда сравнение остается затруднительным.
Однако мы выделяем некоторые случаи, когда сравнение остается затруднительным. Уэст-Эберхард (2003, 2005) экстраполировал различия, сегрегирующие внутри популяций (различие № 2), на различия, возникшие во времени во время эволюции популяции (различие № 1).
Уэст-Эберхард (2003, 2005) экстраполировал различия, сегрегирующие внутри популяций (различие № 2), на различия, возникшие во времени во время эволюции популяции (различие № 1). , или сравнить чувствительность двух разных генотипов к двум разным средам (где фенотипическая вариация становится различием в различии; см., например, Engelman et al., 2009; Томас, 2010).
, или сравнить чувствительность двух разных генотипов к двум разным средам (где фенотипическая вариация становится различием в различии; см., например, Engelman et al., 2009; Томас, 2010). генетические эффекты, поскольку гены и окружающая среда действуют на различных уровнях сложной причинно-следственной связи между генотипами и фенотипами.
генетические эффекты, поскольку гены и окружающая среда действуют на различных уровнях сложной причинно-следственной связи между генотипами и фенотипами. , 2011). Здесь разница в поведении вызвана переключением между присутствием или отсутствием определенного кишечного симбионта. Причиной фенотипических различий является не простое изменение в последовательности ДНК, не простое изменение окружающей среды, не связанное с генетическими изменениями, а переключение между присутствием и отсутствием фактора, который можно рассматривать как фактор окружающей среды, — бактерий, — который содержит ДНК, мутации которой могут также изменить фенотип хозяина.
, 2011). Здесь разница в поведении вызвана переключением между присутствием или отсутствием определенного кишечного симбионта. Причиной фенотипических различий является не простое изменение в последовательности ДНК, не простое изменение окружающей среды, не связанное с генетическими изменениями, а переключение между присутствием и отсутствием фактора, который можно рассматривать как фактор окружающей среды, — бактерий, — который содержит ДНК, мутации которой могут также изменить фенотип хозяина. Вставка мобильного элемента может сбить ген, тогда как делеция иногда может создать новый сайт связывания для активатора транскрипции.На самом деле, эволюционное усиление экспрессии desatF у D. melanogaster произошло за счет серии из трех делеций, каждая из которых создает гексамерный мотив, необходимый для экспрессии desatF (Shirangi et al., 2009).
Вставка мобильного элемента может сбить ген, тогда как делеция иногда может создать новый сайт связывания для активатора транскрипции.На самом деле, эволюционное усиление экспрессии desatF у D. melanogaster произошло за счет серии из трех делеций, каждая из которых создает гексамерный мотив, необходимый для экспрессии desatF (Shirangi et al., 2009). Такая воспроизводимость фенотипического исхода необходима для обеспечения генетической эволюции и адаптации путем естественного отбора (Lewontin, 1974a; Kirschner and Gerhart, 1998).В самом деле, вновь образованный аллель, который будет генерировать еще один фенотип каждый раз, когда он попадает в другой организм, не будет подвергаться естественному отбору. Рассуждение с точки зрения изменчивости, а не рассмотрение аллелей как изолированных объектов, проясняет, что конкуренция происходит между аллелями, которые охватывают один и тот же генетический локус. Естественный отбор воздействует непосредственно на аллельную вариацию, которая устойчиво связана с данной фенотипической вариацией, что и является родством ГП. Таким образом, отношения ГП являются базовой единицей эволюционных изменений, на которые действует естественный отбор.
Такая воспроизводимость фенотипического исхода необходима для обеспечения генетической эволюции и адаптации путем естественного отбора (Lewontin, 1974a; Kirschner and Gerhart, 1998).В самом деле, вновь образованный аллель, который будет генерировать еще один фенотип каждый раз, когда он попадает в другой организм, не будет подвергаться естественному отбору. Рассуждение с точки зрения изменчивости, а не рассмотрение аллелей как изолированных объектов, проясняет, что конкуренция происходит между аллелями, которые охватывают один и тот же генетический локус. Естественный отбор воздействует непосредственно на аллельную вариацию, которая устойчиво связана с данной фенотипической вариацией, что и является родством ГП. Таким образом, отношения ГП являются базовой единицей эволюционных изменений, на которые действует естественный отбор. Другими словами, определенные отношения GP являются таксономически надежными и присутствуют у большого количества видов. Это означает, что генетические и экологические предпосылки оставались относительно постоянными или неоднократно появлялись на протяжении эволюции, что позволяло генетическим локусам генерировать сходные фенотипические изменения в различных таксономических группах.Это важное открытие было совершенно неожиданным около 50 лет назад. Долгое время ожидалось, что сингулярность, наблюдаемая в живом мире, будет отражать сравнимую сингулярность на генетическом уровне, затрагивая несопоставимые и неконсервативные гены, специфичные для каждой линии (Mayr, 1963). Как однажды предположил Майр (1963) в 1963 году: «Многое из того, что было известно о физиологии генов, делает очевидным, что поиск гомологичных генов совершенно бесполезен, за исключением очень близких родственников […]. Поговорка «Многие дороги ведут в Рим» справедлива как в эволюции, так и в повседневных делах» (Mayr, 1963).
Другими словами, определенные отношения GP являются таксономически надежными и присутствуют у большого количества видов. Это означает, что генетические и экологические предпосылки оставались относительно постоянными или неоднократно появлялись на протяжении эволюции, что позволяло генетическим локусам генерировать сходные фенотипические изменения в различных таксономических группах.Это важное открытие было совершенно неожиданным около 50 лет назад. Долгое время ожидалось, что сингулярность, наблюдаемая в живом мире, будет отражать сравнимую сингулярность на генетическом уровне, затрагивая несопоставимые и неконсервативные гены, специфичные для каждой линии (Mayr, 1963). Как однажды предположил Майр (1963) в 1963 году: «Многое из того, что было известно о физиологии генов, делает очевидным, что поиск гомологичных генов совершенно бесполезен, за исключением очень близких родственников […]. Поговорка «Многие дороги ведут в Рим» справедлива как в эволюции, так и в повседневных делах» (Mayr, 1963). Другими словами, ожидалось, что генетические локусы, которые делают человека мужчиной, будут отличаться от тех, которые делают собаку или рыбу. Позднее, в 80–90-х годах, некоторые исследователи предположили, что эволюция происходит за счет мутаций в консервативных генах, кодирующих белок (Romero-Herrera et al., 1978; Perutz, 1983; Stewart et al., 1987; Carroll et al. ., 2005) – но у них было мало экспериментальных данных, подтверждающих их точку зрения (Tautz and Schmid, 1998). Накопившиеся на сегодняшний день данные о мутациях, ответственных за естественную изменчивость, ясно показывают, что разнообразие живых организмов имеет общую генетическую основу как минимум в трех пунктах.Во-первых, сравнительная биология развития показала, что животные имеют общие наборы ключевых регуляторных генов с законсервированными функциями (Wilkins, 2002, 2014; Carroll et al., 2005). Во-вторых, большинство межвидовых различий у животных и растений, для которых хотя бы частично идентифицирована лежащая в основе генетическая основа (154 случая из 160), обусловлено мутациями в гомологичных генах, и очень немногие (6/160) обусловлены новыми генами.
Другими словами, ожидалось, что генетические локусы, которые делают человека мужчиной, будут отличаться от тех, которые делают собаку или рыбу. Позднее, в 80–90-х годах, некоторые исследователи предположили, что эволюция происходит за счет мутаций в консервативных генах, кодирующих белок (Romero-Herrera et al., 1978; Perutz, 1983; Stewart et al., 1987; Carroll et al. ., 2005) – но у них было мало экспериментальных данных, подтверждающих их точку зрения (Tautz and Schmid, 1998). Накопившиеся на сегодняшний день данные о мутациях, ответственных за естественную изменчивость, ясно показывают, что разнообразие живых организмов имеет общую генетическую основу как минимум в трех пунктах.Во-первых, сравнительная биология развития показала, что животные имеют общие наборы ключевых регуляторных генов с законсервированными функциями (Wilkins, 2002, 2014; Carroll et al., 2005). Во-вторых, большинство межвидовых различий у животных и растений, для которых хотя бы частично идентифицирована лежащая в основе генетическая основа (154 случая из 160), обусловлено мутациями в гомологичных генах, и очень немногие (6/160) обусловлены новыми генами. которые, тем не менее, представляют собой дубликаты существующих генов (Martin and Orgogozo, 2013b).В-третьих, было показано, что множественные случаи сходных фенотипических изменений связаны с мутациями одних и тех же гомологичных генов в независимых линиях (таблица 1), иногда на больших филогенетических расстояниях. Например, разница в пигментации между белыми и оранжевыми бенгальскими тиграми недавно была сопоставлена с единственной мутацией в гене белка-транспортера SLC45A2 (Xu et al., 2013), и этот ген также был связан с гипопигментацией глаз, кожи , волосы и перья у людей и кур (Xu et al., 2013; Рисунок 1Е). Более ярким примером является недавняя эволюция устойчивости к токсинам у трех видов, которые разошлись более 500 миллионов лет назад – моллюска, змеи и иглобрюха – посредством замены одной и той же аминокислоты в законсервированном гене (Bricelj et al., 2005). ; Geffeney et al., 2005; Venkatesh et al., 2005; Feldman et al., 2012). Такие поразительные паттерны генетического повторения в настоящее время обнаружены для более чем 100 генов животных и растений (Martin and Orgogozo, 2013b).
которые, тем не менее, представляют собой дубликаты существующих генов (Martin and Orgogozo, 2013b).В-третьих, было показано, что множественные случаи сходных фенотипических изменений связаны с мутациями одних и тех же гомологичных генов в независимых линиях (таблица 1), иногда на больших филогенетических расстояниях. Например, разница в пигментации между белыми и оранжевыми бенгальскими тиграми недавно была сопоставлена с единственной мутацией в гене белка-транспортера SLC45A2 (Xu et al., 2013), и этот ген также был связан с гипопигментацией глаз, кожи , волосы и перья у людей и кур (Xu et al., 2013; Рисунок 1Е). Более ярким примером является недавняя эволюция устойчивости к токсинам у трех видов, которые разошлись более 500 миллионов лет назад – моллюска, змеи и иглобрюха – посредством замены одной и той же аминокислоты в законсервированном гене (Bricelj et al., 2005). ; Geffeney et al., 2005; Venkatesh et al., 2005; Feldman et al., 2012). Такие поразительные паттерны генетического повторения в настоящее время обнаружены для более чем 100 генов животных и растений (Martin and Orgogozo, 2013b). Несмотря на существующие методологические предубеждения в пользу консервативных генов при поиске локусов количественных признаков (Rockman, 2012; Martin and Orgogozo, 2013b), уровень генетического повторения остается поразительным и предполагает, что для эволюции по крайней мере некоторых фенотипических различий требуется относительно мало генетических путей. ведут в Рим (Stern, 2013).В настоящее время не следует удивляться тому, что фрагмент ДНК, связанный со сложной окраской крыльев у одного из видов бабочек Heliconius , обеспечивает сходные крылья и коллективную защиту от одних и тех же хищников при введении в геном других бабочек (Supple et al., 2014). То, что делает собаку собакой или человека человеком, теперь частично объясняется уникальным набором таксономически устойчивых отношений ГП, которые обнаруживаются во многих ветвях линии.
Несмотря на существующие методологические предубеждения в пользу консервативных генов при поиске локусов количественных признаков (Rockman, 2012; Martin and Orgogozo, 2013b), уровень генетического повторения остается поразительным и предполагает, что для эволюции по крайней мере некоторых фенотипических различий требуется относительно мало генетических путей. ведут в Рим (Stern, 2013).В настоящее время не следует удивляться тому, что фрагмент ДНК, связанный со сложной окраской крыльев у одного из видов бабочек Heliconius , обеспечивает сходные крылья и коллективную защиту от одних и тех же хищников при введении в геном других бабочек (Supple et al., 2014). То, что делает собаку собакой или человека человеком, теперь частично объясняется уникальным набором таксономически устойчивых отношений ГП, которые обнаруживаются во многих ветвях линии. У экзотермов температура организма зависит от температуры окружающей среды. Учитывая устрашающее количество условий окружающей среды, которые можно представить, вероятно, невозможно определить, преобладают ли таксономически устойчивые отношения GP или таксономически устойчивые отношения среда-фенотип. Кроме того, вопрос о том, представляют ли таксономически устойчивые отношения GP исключительную и небольшую долю или значительную долю всех отношений GP, является предметом споров.В любом случае, существование таксономически устойчивых взаимосвязей GP теперь ясно и должно быть широко признано исследовательским сообществом.
У экзотермов температура организма зависит от температуры окружающей среды. Учитывая устрашающее количество условий окружающей среды, которые можно представить, вероятно, невозможно определить, преобладают ли таксономически устойчивые отношения GP или таксономически устойчивые отношения среда-фенотип. Кроме того, вопрос о том, представляют ли таксономически устойчивые отношения GP исключительную и небольшую долю или значительную долю всех отношений GP, является предметом споров.В любом случае, существование таксономически устойчивых взаимосвязей GP теперь ясно и должно быть широко признано исследовательским сообществом.
 Предсказуемость GP уже используется для идентификации штаммов, которые развили устойчивость к различным стратегиям борьбы с вредителями, причем крайние случаи нацелены на толерантность к противомалярийным препаратам у паразитов Plasmodium (Manske et al., 2012), устойчивость бактерий и дрожжей к антибиотикам (Fischbach, 2009; MacCallum et al., 2010) или, что еще более драматично, антропогенная эволюция устойчивости к инсектицидам у различных когорт насекомых, независимо от их статуса вредителя (Ffrench-Constant и др., 2004; Мартин и Оргогозо, 2013b).
Предсказуемость GP уже используется для идентификации штаммов, которые развили устойчивость к различным стратегиям борьбы с вредителями, причем крайние случаи нацелены на толерантность к противомалярийным препаратам у паразитов Plasmodium (Manske et al., 2012), устойчивость бактерий и дрожжей к антибиотикам (Fischbach, 2009; MacCallum et al., 2010) или, что еще более драматично, антропогенная эволюция устойчивости к инсектицидам у различных когорт насекомых, независимо от их статуса вредителя (Ffrench-Constant и др., 2004; Мартин и Оргогозо, 2013b). , 2013), что расширило использование модельных организмов в качестве моделей болезней.
, 2013), что расширило использование модельных организмов в качестве моделей болезней. , 2013). В таких случаях интуитивно понятно сгруппировать такие генетически связанные сайты вместе, поскольку все они влияют на один и тот же тип фенотипического признака.
, 2013). В таких случаях интуитивно понятно сгруппировать такие генетически связанные сайты вместе, поскольку все они влияют на один и тот же тип фенотипического признака. Это явно относится к сигнальным молекулам TGF-β BMP15, GDF9 и рецептору TGF-β BMPR1B, которые неоднократно связывались с изменениями функции яичников у людей и пород домашних овец (обзор в Luong et al., 2011).
Это явно относится к сигнальным молекулам TGF-β BMP15, GDF9 и рецептору TGF-β BMPR1B, которые неоднократно связывались с изменениями функции яичников у людей и пород домашних овец (обзор в Luong et al., 2011). Этот несколько пограничный пример иллюстрирует проблему включения широко распространенного сравнительного мышления в наше глобальное понимание биологии. Имеет ли отношение мутация в мышиной модели к заболеванию человека? Можем ли мы считать, что фенотип мыши подобен состоянию человека, если его генетическая основа иная? Мы и другие прогнозируем, что поиск ортологичных фенотипов, или «фенологов» (McGary et al., 2010), станет главной задачей современной генетики и потребует плодотворного союза между прикладной и эволюционной биологией.
Этот несколько пограничный пример иллюстрирует проблему включения широко распространенного сравнительного мышления в наше глобальное понимание биологии. Имеет ли отношение мутация в мышиной модели к заболеванию человека? Можем ли мы считать, что фенотип мыши подобен состоянию человека, если его генетическая основа иная? Мы и другие прогнозируем, что поиск ортологичных фенотипов, или «фенологов» (McGary et al., 2010), станет главной задачей современной генетики и потребует плодотворного союза между прикладной и эволюционной биологией.
 (2014). Вариант нейропептидного рецептора npr-1 является основной детерминантой роста и физиологии Caenorhabditis elegans . Генетика PLoS. 10:e1004156. doi: 10.1371/journal.pgen.1004156
(2014). Вариант нейропептидного рецептора npr-1 является основной детерминантой роста и физиологии Caenorhabditis elegans . Генетика PLoS. 10:e1004156. doi: 10.1371/journal.pgen.1004156 Проц.Натл. акад. науч. США 108, 16050–16055. doi: 10.1073/pnas.1102999108
Проц.Натл. акад. науч. США 108, 16050–16055. doi: 10.1073/pnas.1102999108 дои: 10.1515/9781400841653
дои: 10.1515/9781400841653 PLoS Биол. 5:e96. doi: 10.1371/journal.pbio.0050096
PLoS Биол. 5:e96. doi: 10.1371/journal.pbio.0050096 Природа 474, 598–603. doi: 10.1038/nature10200
Природа 474, 598–603. doi: 10.1038/nature10200
 , Хэтчер, Дж. Л., и Дайресон, Э. Г. (2000). Генетический и онтогенетический анализ различий в пигментации брюшка у разных видов подгруппы Drosophila dunni . Эволюция. Междунар. Дж. Орг. Эвол. 54, 2057–2071. doi: 10.1111/j.0014-3820.2000.tb01249.x
, Хэтчер, Дж. Л., и Дайресон, Э. Г. (2000). Генетический и онтогенетический анализ различий в пигментации брюшка у разных видов подгруппы Drosophila dunni . Эволюция. Междунар. Дж. Орг. Эвол. 54, 2057–2071. doi: 10.1111/j.0014-3820.2000.tb01249.x , Ван С., Тан Дж., Жербо П., Варк А., Тан Л. и соавт. (2013). Моделирование недавней эволюции человека у мышей путем экспрессии выбранного варианта EDAR. Сотовый 152, 691–702. дои: 10.1016/j.cell.2013.01.016
, Ван С., Тан Дж., Жербо П., Варк А., Тан Л. и соавт. (2013). Моделирование недавней эволюции человека у мышей путем экспрессии выбранного варианта EDAR. Сотовый 152, 691–702. дои: 10.1016/j.cell.2013.01.016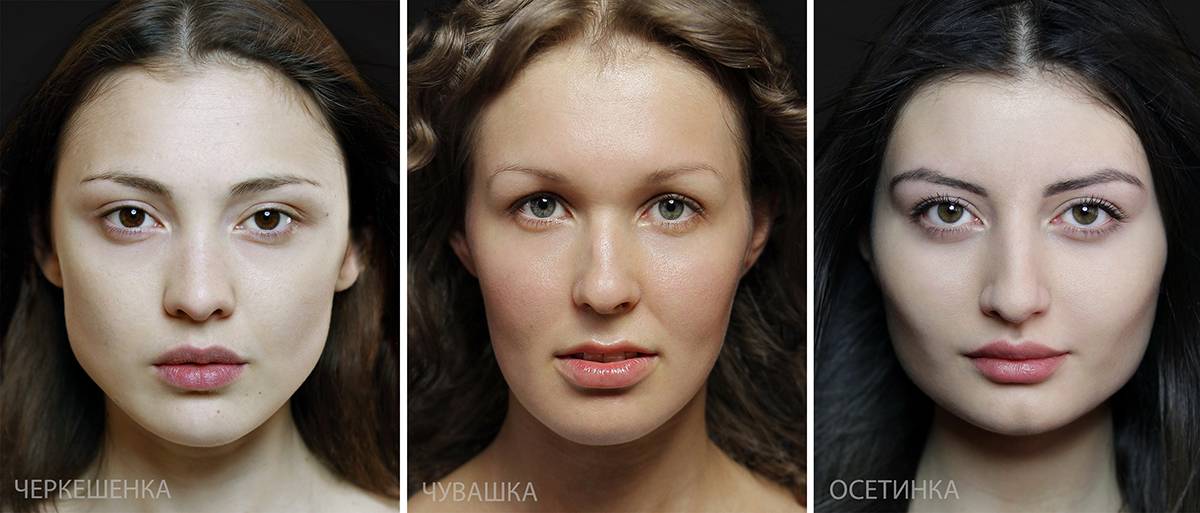 Натл. акад. науч. США 95, 8420–8427. doi: 10.1073/pnas.95.15.8420
Натл. акад. науч. США 95, 8420–8427. doi: 10.1073/pnas.95.15.8420 0504210102
0504210102 Междунар. соц. Твин Стад. 14, 408–416. doi: 10.1375/twin.14.5.408
Междунар. соц. Твин Стад. 14, 408–416. doi: 10.1375/twin.14.5.408 и Оргогозо В. (2013b). Локусы повторной эволюции: каталог генетических очагов фенотипической изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 67, 1235–1250. doi: 10.1111/evo.12081
и Оргогозо В. (2013b). Локусы повторной эволюции: каталог генетических очагов фенотипической изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 67, 1235–1250. doi: 10.1111/evo.12081 , Вебер Р. Э., Фаго А., Морияма Х. и Сторц Дж. Ф. (2013). Эпистаз среди адаптивных мутаций гемоглобина оленьих мышей. Наука 340, 1324–1327. doi: 10.1126/science.1236862
, Вебер Р. Э., Фаго А., Морияма Х. и Сторц Дж. Ф. (2013). Эпистаз среди адаптивных мутаций гемоглобина оленьих мышей. Наука 340, 1324–1327. doi: 10.1126/science.1236862 Х., Лин, Ю. Р., Ли, З., Шерц, К. Ф., Добли, Дж. Ф., Пинсон, С. Р., и соавт. (1995). Конвергентное одомашнивание злаковых культур путем независимых мутаций в соответствующих генетических локусах. Наука 269, 1714–1718. doi: 10.1126/наука.269.5231.1714
Х., Лин, Ю. Р., Ли, З., Шерц, К. Ф., Добли, Дж. Ф., Пинсон, С. Р., и соавт. (1995). Конвергентное одомашнивание злаковых культур путем независимых мутаций в соответствующих генетических локусах. Наука 269, 1714–1718. doi: 10.1126/наука.269.5231.1714 А., Саломоне, Дж. Р., Тейси, Д. Дж., Камино, Э. М., Дэвис, К. А., Ребейс, М., и соавт. (2013). Рекуррентная модификация консервативного цис-регуляторного элемента лежит в основе разнообразия пигментации плодовых мушек. PLoS Genet 9:e1003740. doi: 10.1371/journal.pgen.1003740
А., Саломоне, Дж. Р., Тейси, Д. Дж., Камино, Э. М., Дэвис, К. А., Ребейс, М., и соавт. (2013). Рекуррентная модификация консервативного цис-регуляторного элемента лежит в основе разнообразия пигментации плодовых мушек. PLoS Genet 9:e1003740. doi: 10.1371/journal.pgen.1003740 Falk and HJ Rheinberger (Cambridge, NY: Cambridge University Press), 26–39. . дои: 10.1017/CBO9780511527296.004
Falk and HJ Rheinberger (Cambridge, NY: Cambridge University Press), 26–39. . дои: 10.1017/CBO9780511527296.004 Эволюционная биология развития и проблема изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 54, 1079–1091. doi: 10.1111/j.0014-3820.2000.tb00544.x
Эволюционная биология развития и проблема изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 54, 1079–1091. doi: 10.1111/j.0014-3820.2000.tb00544.x Филос. Транс. Р. Соц. Лонд. Б. биол. науч. 353, 231–240. doi: 10.1098/rstb.1998.0205
Филос. Транс. Р. Соц. Лонд. Б. биол. науч. 353, 231–240. doi: 10.1098/rstb.1998.0205 Принстон, Нью-Джерси: Издательство Принстонского университета. дои: 10.1515/9781400851461
Принстон, Нью-Джерси: Издательство Принстонского университета. дои: 10.1515/9781400851461 Оксфорд: Издательство Оксфордского университета.
Оксфорд: Издательство Оксфордского университета. Однако значительная проблема заключается в том, как люди могут сопоставлять диапазон визуальных фонов, на которых их часто видят. Большинство типов камуфляжа связано с взаимодействием между окружающей средой и фенотипом животного, при этом внешний вид должен в большей или меньшей степени напоминать фон, на котором наблюдается животное. Большинство мест обитания не однородны, а демонстрируют значительное разнообразие цвета, яркости и рисунка.Одно из решений этой проблемы включает в себя отбор, благоприятствующий эволюции генетических полиморфизмов внешнего вида, когда особи попадают в одну из множества дискретных морфов. Такие полиморфизмы и их генетическая основа хорошо изучены у мышей и рептилий (Nachman et al., 2003; Rosenblum et al., 2004; Rosenblum, 2006).
Однако значительная проблема заключается в том, как люди могут сопоставлять диапазон визуальных фонов, на которых их часто видят. Большинство типов камуфляжа связано с взаимодействием между окружающей средой и фенотипом животного, при этом внешний вид должен в большей или меньшей степени напоминать фон, на котором наблюдается животное. Большинство мест обитания не однородны, а демонстрируют значительное разнообразие цвета, яркости и рисунка.Одно из решений этой проблемы включает в себя отбор, благоприятствующий эволюции генетических полиморфизмов внешнего вида, когда особи попадают в одну из множества дискретных морфов. Такие полиморфизмы и их генетическая основа хорошо изучены у мышей и рептилий (Nachman et al., 2003; Rosenblum et al., 2004; Rosenblum, 2006).
 Что касается камуфляжа, то большинство исследований было сосредоточено на относительно небольшом количестве видов, в частности на тех, которые способны к чрезвычайно быстрым изменениям (секунды), особенно на головоногих (Hanlon et al., 2009; Цзяо и др., 2011 г.; Зилински и Йонсен, 2011). Хотя эти исследования показали многое о том, как изменение цвета можно использовать для соответствия различным средам, и связанную с этим визуальную обратную связь, изменение цвета с такой скоростью, вероятно, будет нетипичным по своей природе, особенно за пределами головоногих моллюсков и рыб. Вместо этого большинство животных, вероятно, меняются в течение нескольких часов, дней, недель и месяцев (Umbers et al., 2014). Это может включать модификации, происходящие в результате физиологического изменения цвета, которые часто связаны с сокращением и диспергированием окрашенного пигмента в хроматофорных клетках, долгосрочные изменения, связанные с морфологией клеточного распределения и синтеза пигмента, а также в процессе развития (Stuart-Fox and Moussalli, 2009; Амберс и др.
Что касается камуфляжа, то большинство исследований было сосредоточено на относительно небольшом количестве видов, в частности на тех, которые способны к чрезвычайно быстрым изменениям (секунды), особенно на головоногих (Hanlon et al., 2009; Цзяо и др., 2011 г.; Зилински и Йонсен, 2011). Хотя эти исследования показали многое о том, как изменение цвета можно использовать для соответствия различным средам, и связанную с этим визуальную обратную связь, изменение цвета с такой скоростью, вероятно, будет нетипичным по своей природе, особенно за пределами головоногих моллюсков и рыб. Вместо этого большинство животных, вероятно, меняются в течение нескольких часов, дней, недель и месяцев (Umbers et al., 2014). Это может включать модификации, происходящие в результате физиологического изменения цвета, которые часто связаны с сокращением и диспергированием окрашенного пигмента в хроматофорных клетках, долгосрочные изменения, связанные с морфологией клеточного распределения и синтеза пигмента, а также в процессе развития (Stuart-Fox and Moussalli, 2009; Амберс и др. , 2014). Большинство исследований в основном сосредоточены на физиологических процессах, не всегда рассматривают функциональные и экологические проблемы, и остается множество вопросов о механизмах и адаптивном значении изменения цвета.
, 2014). Большинство исследований в основном сосредоточены на физиологических процессах, не всегда рассматривают функциональные и экологические проблемы, и остается множество вопросов о механизмах и адаптивном значении изменения цвета. Прошлые работы были сосредоточены, в частности, на крабах-скрипачах, Uca .Некоторые виды имеют циркадный дневной и ночной ритм изменения окраски (преимущественно яркости), при котором особи часто становятся светлее ночью и темнее днем (Аткинс, 1926; Абрамовиц, 1937; Браун и Уэбб, 1948). Это может быть связано с изменением дисперсии черного и белого пигментов в хроматофорных клетках (Abramowitz, 1937; Brown, Webb, 1948; Fingerman, 1955, 1956; Fingerman, Yamamoto, 1967; Rao et al., 1967) и наиболее вероятно. выполнять функцию защиты крабов от интенсивного ультрафиолетового излучения (Darnell, 2012).Некоторые крабы-скрипачи также демонстрируют более высокие уровни дисперсии черного и белого пигмента на темном и светлом фоне соответственно (Brown and Sandeen, 1948; Rao et al., 1967), предположительно для маскировки. В целом, несмотря на значительную работу по изменению цвета крабов Uca , большинство из них не касалось маскировки и не фокусировалось на эколого-эволюционном значении изменения цвета.
Прошлые работы были сосредоточены, в частности, на крабах-скрипачах, Uca .Некоторые виды имеют циркадный дневной и ночной ритм изменения окраски (преимущественно яркости), при котором особи часто становятся светлее ночью и темнее днем (Аткинс, 1926; Абрамовиц, 1937; Браун и Уэбб, 1948). Это может быть связано с изменением дисперсии черного и белого пигментов в хроматофорных клетках (Abramowitz, 1937; Brown, Webb, 1948; Fingerman, 1955, 1956; Fingerman, Yamamoto, 1967; Rao et al., 1967) и наиболее вероятно. выполнять функцию защиты крабов от интенсивного ультрафиолетового излучения (Darnell, 2012).Некоторые крабы-скрипачи также демонстрируют более высокие уровни дисперсии черного и белого пигмента на темном и светлом фоне соответственно (Brown and Sandeen, 1948; Rao et al., 1967), предположительно для маскировки. В целом, несмотря на значительную работу по изменению цвета крабов Uca , большинство из них не касалось маскировки и не фокусировалось на эколого-эволюционном значении изменения цвета. Кроме того, большая часть работы, либо в течение дневных и лунных циклов, либо на разных фонах, включала оценку изменения цвета с использованием индекса дисперсии пигмента, который показывает, насколько рассредоточенные типы пигмента кажутся человеческому глазу под микроскопом.В сравнительно небольшом количестве исследований объективно оценивали изменение окраски крабов (но см. Hemmi et al., 2006; Detto et al., 2008; Darnell, 2012) и редко их маскировку (Stevens et al., 2013, 2014a; Russell and Dierssen, 2015). ).
Кроме того, большая часть работы, либо в течение дневных и лунных циклов, либо на разных фонах, включала оценку изменения цвета с использованием индекса дисперсии пигмента, который показывает, насколько рассредоточенные типы пигмента кажутся человеческому глазу под микроскопом.В сравнительно небольшом количестве исследований объективно оценивали изменение окраски крабов (но см. Hemmi et al., 2006; Detto et al., 2008; Darnell, 2012) и редко их маскировку (Stevens et al., 2013, 2014a; Russell and Dierssen, 2015). ). 1). Эти изменения, по-видимому, позволяют им лучше соответствовать цвету и яркости песчаного пляжа в течение дня и, возможно, сливаться с тенями ночью. Кроме того, как и Uca , крабы становятся светлее на белом фоне и темнее на черном фоне, что потенциально еще больше улучшает их камуфляж.Изменения яркости не происходит, когда крабов просто помещают в темноту. Эти эксперименты показывают, как изменение цвета потенциально можно использовать для модуляции маскировки в течение нескольких часов, хотя в исследовании не моделировался внешний вид или маскировка крабов-призраков для зрения хищника. В другой работе изучалась окраска и камуфляж крабов, обнаруженных на матах Sargassum , которые, по-видимому, также демонстрируют заметные изменения внешнего вида в зависимости от субстрата (Russell and Dierssen, 2015). В этой работе моделировался внешний вид и маскировка крабов для рыб и птичьего зрения, при этом камуфляж крабов лучше противостоял дихроматической системе рыб.
1). Эти изменения, по-видимому, позволяют им лучше соответствовать цвету и яркости песчаного пляжа в течение дня и, возможно, сливаться с тенями ночью. Кроме того, как и Uca , крабы становятся светлее на белом фоне и темнее на черном фоне, что потенциально еще больше улучшает их камуфляж.Изменения яркости не происходит, когда крабов просто помещают в темноту. Эти эксперименты показывают, как изменение цвета потенциально можно использовать для модуляции маскировки в течение нескольких часов, хотя в исследовании не моделировался внешний вид или маскировка крабов-призраков для зрения хищника. В другой работе изучалась окраска и камуфляж крабов, обнаруженных на матах Sargassum , которые, по-видимому, также демонстрируют заметные изменения внешнего вида в зависимости от субстрата (Russell and Dierssen, 2015). В этой работе моделировался внешний вид и маскировка крабов для рыб и птичьего зрения, при этом камуфляж крабов лучше противостоял дихроматической системе рыб. 1. Левое изображение показывает изменение яркости двух береговых крабов ( Carcinus maenas ) при помещении на черный (слева) и белый (справа) фон на 2 часа . Из Stevens et al. (2014а). На правом изображении показан рогатый краб-призрак днем со светло-желтой окраской (вверху) и ночью с темно-серой окраской (внизу).
1. Левое изображение показывает изменение яркости двух береговых крабов ( Carcinus maenas ) при помещении на черный (слева) и белый (справа) фон на 2 часа . Из Stevens et al. (2014а). На правом изображении показан рогатый краб-призрак днем со светло-желтой окраской (вверху) и ночью с темно-серой окраской (внизу). В наших экспериментах мы проверили молодь крабов на изменение яркости на черном или белом фоне и на изменение цвета на красном или зеленом фоне, а также смоделировали степень изменения цвета и яркости (воспринимаемой легкости) особей в зрительной системе человека. птичий хищник.Мы обнаружили, что крабы могут менять свою яркость, становясь темнее на черном фоне и светлее на белом фоне (рис. 1). Хотя существовали значительные индивидуальные различия, и изменения часто не были значительными, у некоторых людей они были достаточно значительными, чтобы привести к лучшему совпадению фона с моделью визуального различения. Будет ли это равносильно снижению риска хищничества, требует проверки с помощью экспериментов по хищничеству / обнаружению. Напротив, мы не обнаружили изменений в цвете на зеленом и красном фоне, по крайней мере, в той степени, в которой это могло бы повлиять на камуфляж.
В наших экспериментах мы проверили молодь крабов на изменение яркости на черном или белом фоне и на изменение цвета на красном или зеленом фоне, а также смоделировали степень изменения цвета и яркости (воспринимаемой легкости) особей в зрительной системе человека. птичий хищник.Мы обнаружили, что крабы могут менять свою яркость, становясь темнее на черном фоне и светлее на белом фоне (рис. 1). Хотя существовали значительные индивидуальные различия, и изменения часто не были значительными, у некоторых людей они были достаточно значительными, чтобы привести к лучшему совпадению фона с моделью визуального различения. Будет ли это равносильно снижению риска хищничества, требует проверки с помощью экспериментов по хищничеству / обнаружению. Напротив, мы не обнаружили изменений в цвете на зеленом и красном фоне, по крайней мере, в той степени, в которой это могло бы повлиять на камуфляж. Во-первых, появляется все больше свидетельств того, что у молодых особей могут происходить некоторые изменения яркости, которые могут улучшить маскировку (Powell, 1962b; Stevens et al., 2014a). Кроме того, они, весьма вероятно, претерпевают существенные изменения внешнего вида за счет фенотипической пластичности во время линьки (Todd et al., 2006; Stevens et al., 2014b; Jensen and Egnotovich, 2015; рис. 2).Действительно, паттерны передачи сигналов у крабов-скрипачей могут претерпевать существенные изменения между линьками (Detto et al., 2008). Во-вторых, береговые крабы — распространенный и широко распространенный вид, встречающийся во многих местах обитания, потенциально требующий разного внешнего вида для обеспечения маскировки в каждом из них. В соответствии с этим крабы из каменистых, илистых и мидиевых местообитаний демонстрируют различия как в цвете, так и в рисунке (Todd et al., 2006, 2012; Stevens et al., 2014b; рисунок 3). Тем не менее, стоит отметить, что, несмотря на ряд исследований, изучающих ассоциации фенотип-окружающая среда , которые, вероятно, способствуют камуфляжу (для целого ряда таксонов животных), только одно исследование песчаных блох непосредственно тестировало камуфляж (напр.
Во-первых, появляется все больше свидетельств того, что у молодых особей могут происходить некоторые изменения яркости, которые могут улучшить маскировку (Powell, 1962b; Stevens et al., 2014a). Кроме того, они, весьма вероятно, претерпевают существенные изменения внешнего вида за счет фенотипической пластичности во время линьки (Todd et al., 2006; Stevens et al., 2014b; Jensen and Egnotovich, 2015; рис. 2).Действительно, паттерны передачи сигналов у крабов-скрипачей могут претерпевать существенные изменения между линьками (Detto et al., 2008). Во-вторых, береговые крабы — распространенный и широко распространенный вид, встречающийся во многих местах обитания, потенциально требующий разного внешнего вида для обеспечения маскировки в каждом из них. В соответствии с этим крабы из каменистых, илистых и мидиевых местообитаний демонстрируют различия как в цвете, так и в рисунке (Todd et al., 2006, 2012; Stevens et al., 2014b; рисунок 3). Тем не менее, стоит отметить, что, несмотря на ряд исследований, изучающих ассоциации фенотип-окружающая среда , которые, вероятно, способствуют камуфляжу (для целого ряда таксонов животных), только одно исследование песчаных блох непосредственно тестировало камуфляж (напр. г., фенотип-среда соответствует ; Стивенс и др., 2015). В-третьих, береговые крабы сильно различаются по внешнему виду даже в одном и том же месте, по-видимому, с широким спектром тактик маскировки от совпадения с фоном до разрушительной окраски (Hogarth, 1978; Stevens et al., 2014b; рис. 4). Уровень разнообразия связан с типом среды обитания / местом, где они живут. Это открытие может позволить проверить, насколько высокое внутривидовое разнообразие создается и поддерживается. В-четвертых, их биология и крепкий характер делают их идеальным видом для проведения экспериментов и определения механизмов изменения цвета.Наконец, за пределами своего естественного европейского ареала они высоко инвазивны по всему миру (один из «100 лучших» инвазивных видов в мире), а это означает, что они представляют собой ценную систему для изучения биологии инвазии (Darling et al., 2008; McGaw et al. ., 2011).
г., фенотип-среда соответствует ; Стивенс и др., 2015). В-третьих, береговые крабы сильно различаются по внешнему виду даже в одном и том же месте, по-видимому, с широким спектром тактик маскировки от совпадения с фоном до разрушительной окраски (Hogarth, 1978; Stevens et al., 2014b; рис. 4). Уровень разнообразия связан с типом среды обитания / местом, где они живут. Это открытие может позволить проверить, насколько высокое внутривидовое разнообразие создается и поддерживается. В-четвертых, их биология и крепкий характер делают их идеальным видом для проведения экспериментов и определения механизмов изменения цвета.Наконец, за пределами своего естественного европейского ареала они высоко инвазивны по всему миру (один из «100 лучших» инвазивных видов в мире), а это означает, что они представляют собой ценную систему для изучения биологии инвазии (Darling et al., 2008; McGaw et al. ., 2011). Как и многим другим животным, это, по-видимому, позволяет им соответствовать внешнему виду фона для маскировки.
Как и многим другим животным, это, по-видимому, позволяет им соответствовать внешнему виду фона для маскировки. (2014б).
(2014б). , 2014). Конечно, изменение цвета для маскировки было зарегистрировано у многих групп животных, включая чешуекрылых, ракообразных, головоногих, рептилий, амфибий, рыб и многих других. У видов Uca циркадные изменения могут обеспечить дневную защиту от ультрафиолетового излучения в более низких широтах. Однако защита от ультрафиолетовых лучей менее вероятна у прибрежных крабов с более умеренным климатом, особенно с учетом северного распространения их ареала. Напротив, причиной может быть терморегуляция (на мелководье или на открытом воздухе) в течение дня.У крабов-призраков, которые становятся светлее в течение дня, функция, по-видимому, заключается в маскировке, поскольку это изменение улучшает уровень соответствия пляжному субстрату. Для всех трех видов крабов кратковременные изменения в соответствии с фоновой яркостью, по-видимому, служат для маскировки. Обратите внимание, однако, что было проведено несколько прямых экспериментальных испытаний этих предложенных функций (см. Ниже).
, 2014). Конечно, изменение цвета для маскировки было зарегистрировано у многих групп животных, включая чешуекрылых, ракообразных, головоногих, рептилий, амфибий, рыб и многих других. У видов Uca циркадные изменения могут обеспечить дневную защиту от ультрафиолетового излучения в более низких широтах. Однако защита от ультрафиолетовых лучей менее вероятна у прибрежных крабов с более умеренным климатом, особенно с учетом северного распространения их ареала. Напротив, причиной может быть терморегуляция (на мелководье или на открытом воздухе) в течение дня.У крабов-призраков, которые становятся светлее в течение дня, функция, по-видимому, заключается в маскировке, поскольку это изменение улучшает уровень соответствия пляжному субстрату. Для всех трех видов крабов кратковременные изменения в соответствии с фоновой яркостью, по-видимому, служат для маскировки. Обратите внимание, однако, что было проведено несколько прямых экспериментальных испытаний этих предложенных функций (см. Ниже).
 , 2006; Stevens et al., 2014b, 2015; Hultgren and Mittelstaedt, 2015; Jensen and Egnotovich, 2015; Russell and Dierssen, 2015; Duarte and Flores, 2016). Это может быть особенно ценным для видов с высокой степенью расселения и планктонных личиночных стадий, таких как многие ракообразные, поскольку может быть неопределенность в отношении того, где будет оседать молодь и какова визуальная среда.Еще одной особенностью многих из этих видов является то, что они имеют высокий уровень внутривидового разнообразия по окраске и рисунку (рис. 4, 5) даже в одной и той же местности (Keeble and Gamble, 1899; Gamble and Keeble, 1900; Todd et al., 2006). ; Stevens et al., 2014b; Carvalho-Batista et al., 2015; Hultgren and Mittelstaedt, 2015; Jensen and Egnotovich, 2015; Duarte and Flores, 2016). Причины этого пока во многом неясны. У таких видов, как береговой краб, это может позволить сопоставить ряд различных типов пятен в неоднородных средах обитания или предотвратить формирование изображений поиска хищника (Stevens et al.
, 2006; Stevens et al., 2014b, 2015; Hultgren and Mittelstaedt, 2015; Jensen and Egnotovich, 2015; Russell and Dierssen, 2015; Duarte and Flores, 2016). Это может быть особенно ценным для видов с высокой степенью расселения и планктонных личиночных стадий, таких как многие ракообразные, поскольку может быть неопределенность в отношении того, где будет оседать молодь и какова визуальная среда.Еще одной особенностью многих из этих видов является то, что они имеют высокий уровень внутривидового разнообразия по окраске и рисунку (рис. 4, 5) даже в одной и той же местности (Keeble and Gamble, 1899; Gamble and Keeble, 1900; Todd et al., 2006). ; Stevens et al., 2014b; Carvalho-Batista et al., 2015; Hultgren and Mittelstaedt, 2015; Jensen and Egnotovich, 2015; Duarte and Flores, 2016). Причины этого пока во многом неясны. У таких видов, как береговой краб, это может позволить сопоставить ряд различных типов пятен в неоднородных средах обитания или предотвратить формирование изображений поиска хищника (Stevens et al. , 2014б). Они также могут представлять различные типы стратегий маскировки, такие как совпадение фона или разрушительная окраска. Еще один в значительной степени нерешенный вопрос заключается в том, почему многие крабы сильно различаются по внешнему виду среди особей в молодости, но претерпевают онтогенетические изменения по мере своего роста (рис. 3). У многих видов взрослые особи становятся менее узорчатыми, часто более темными и, по-видимому, менее загадочными (Palma and Steneck, 2001; Todd et al., 2006, 2009; Stevens et al., 2014b; Carvalho-Batista et al., 2015; Jensen and Эгнотович, 2015; Рассел и Дирсен, 2015).Два возможных объяснения заключаются в том, что более старые/более крупные крабы перемещаются в другие типы местообитаний, требуя другого типа маскировки, или что по мере того, как они становятся больше, они становятся менее восприимчивыми к хищникам (Todd et al., 2009). У береговых крабов молодь, вероятно, очень уязвима как для рыб, так и для хищников-птиц, тогда как взрослые особи более устойчивы к нападению со стороны мелких и средних рыб.
, 2014б). Они также могут представлять различные типы стратегий маскировки, такие как совпадение фона или разрушительная окраска. Еще один в значительной степени нерешенный вопрос заключается в том, почему многие крабы сильно различаются по внешнему виду среди особей в молодости, но претерпевают онтогенетические изменения по мере своего роста (рис. 3). У многих видов взрослые особи становятся менее узорчатыми, часто более темными и, по-видимому, менее загадочными (Palma and Steneck, 2001; Todd et al., 2006, 2009; Stevens et al., 2014b; Carvalho-Batista et al., 2015; Jensen and Эгнотович, 2015; Рассел и Дирсен, 2015).Два возможных объяснения заключаются в том, что более старые/более крупные крабы перемещаются в другие типы местообитаний, требуя другого типа маскировки, или что по мере того, как они становятся больше, они становятся менее восприимчивыми к хищникам (Todd et al., 2009). У береговых крабов молодь, вероятно, очень уязвима как для рыб, так и для хищников-птиц, тогда как взрослые особи более устойчивы к нападению со стороны мелких и средних рыб. Тем не менее функция этого паттерна онтогенетических изменений несколько загадочна. Это может помочь нам узнать, как маскировка работает и взаимодействует с размером тела, а также о компромиссах с другими функциями.
Тем не менее функция этого паттерна онтогенетических изменений несколько загадочна. Это может помочь нам узнать, как маскировка работает и взаимодействует с размером тела, а также о компромиссах с другими функциями. Стюарт-Фокс и Муссалли, 2009 г.). Опять же, у крабов была проделана большая работа по физиологическим изменениям.
Стюарт-Фокс и Муссалли, 2009 г.). Опять же, у крабов была проделана большая работа по физиологическим изменениям. различные временные масштабы (особенно онтогенетические изменения) и не связывали это с количественной оценкой изменений общей окраски.Кроме того, то, что происходит во время линьки у ракообразных, насколько мне известно, не исследовано. В идеале будущая работа должна сочетать современные достижения в идентификации пигментных клеток и типов (например, Saenko et al., 2013) с количественной оценкой и визуальным моделированием изменений общего внешнего вида. Вдобавок ко всему, роль генетики и молекулярных путей в потенциале и функционировании изменения цвета пока неизвестна.
различные временные масштабы (особенно онтогенетические изменения) и не связывали это с количественной оценкой изменений общей окраски.Кроме того, то, что происходит во время линьки у ракообразных, насколько мне известно, не исследовано. В идеале будущая работа должна сочетать современные достижения в идентификации пигментных клеток и типов (например, Saenko et al., 2013) с количественной оценкой и визуальным моделированием изменений общего внешнего вида. Вдобавок ко всему, роль генетики и молекулярных путей в потенциале и функционировании изменения цвета пока неизвестна. и Гэмбл, 1899 г.; Гэмбл и Кибл, 1900 г.; Самнер и Киз, 1929 г.). Это согласуется с работой, показывающей, что, например, крабы-призраки не просто становятся темнее, если их поместить в темноту (Stevens et al., 2013). Такая информация может быть получена при различном излучении света, падающего на разные части глаза (Браун и Сандин, 1948). Что касается цвета, у животных может быть либо система цветового зрения, способная различать разные цвета фона, либо, как в случае с каракатицей, они могут быть дальтониками и потенциально использовать эмпирическое правило о том, как различная яркость и объекты преобразуются в цвет. , в сочетании с цветными пигментами, близкими к природным субстратам (Mäthger et al., 2006, 2008). В качестве альтернативы или в дополнение каракатица может в первую очередь соответствовать яркости и рисунку фона, если это наиболее важно для облегчения маскировки. Хотя эти идеи кажутся логичными и интуитивными, очень немногие эксперименты проверяли эти предположения. Наша работа с береговыми крабами (Stevens et al.
и Гэмбл, 1899 г.; Гэмбл и Кибл, 1900 г.; Самнер и Киз, 1929 г.). Это согласуется с работой, показывающей, что, например, крабы-призраки не просто становятся темнее, если их поместить в темноту (Stevens et al., 2013). Такая информация может быть получена при различном излучении света, падающего на разные части глаза (Браун и Сандин, 1948). Что касается цвета, у животных может быть либо система цветового зрения, способная различать разные цвета фона, либо, как в случае с каракатицей, они могут быть дальтониками и потенциально использовать эмпирическое правило о том, как различная яркость и объекты преобразуются в цвет. , в сочетании с цветными пигментами, близкими к природным субстратам (Mäthger et al., 2006, 2008). В качестве альтернативы или в дополнение каракатица может в первую очередь соответствовать яркости и рисунку фона, если это наиболее важно для облегчения маскировки. Хотя эти идеи кажутся логичными и интуитивными, очень немногие эксперименты проверяли эти предположения. Наша работа с береговыми крабами (Stevens et al. , 2014a) не выявила признаков изменения цвета на красном и зеленом фоне. Это может произойти, если крабы не могут определить разницу между этими цветами (или при отсутствии различий в яркости, если крабы используют ахроматические механизмы для сопоставления цветов).Кажется вероятным, что береговые крабы могут обнаруживать различия между относительно коротковолновым («синим») и длинноволновым («красным» или «желтым») светом (Martin and Mote, 1982), но неясно, могут ли они различать длинноволновый и средневолновой свет. («зеленый», «красный» и «желтый»). Однако более долгосрочные изменения цвета все же происходят (рис. 2), поэтому, скорее всего, проблема в том, что временной масштаб наших экспериментов был очень коротким.
, 2014a) не выявила признаков изменения цвета на красном и зеленом фоне. Это может произойти, если крабы не могут определить разницу между этими цветами (или при отсутствии различий в яркости, если крабы используют ахроматические механизмы для сопоставления цветов).Кажется вероятным, что береговые крабы могут обнаруживать различия между относительно коротковолновым («синим») и длинноволновым («красным» или «желтым») светом (Martin and Mote, 1982), но неясно, могут ли они различать длинноволновый и средневолновой свет. («зеленый», «красный» и «желтый»). Однако более долгосрочные изменения цвета все же происходят (рис. 2), поэтому, скорее всего, проблема в том, что временной масштаб наших экспериментов был очень коротким. Однако исследования крабов Рак с удалением либо целых стебельков глаз, либо только комплекса синусовых желез (источник гормонов, диспергирующих пигмент, на стебельке глаз), либо разрезания зрительных нервов позволяют предположить, что визуальная информация через зрительный нерв является ключом к контролю дисперсии пигмента, а не наличие только синусовых желез (Shibley, 1968).То есть визуальные сигналы, проходящие к зрительной доле, кажутся решающими в контроле ответов (предположительно, через другие источники гормонов, не связанные с стебельками глаз). Поэтому необходимо проверить, как визуальная информация и роль гормонов связаны у крабов и других животных, чтобы понять пути, которыми контролируются изменения.
Однако исследования крабов Рак с удалением либо целых стебельков глаз, либо только комплекса синусовых желез (источник гормонов, диспергирующих пигмент, на стебельке глаз), либо разрезания зрительных нервов позволяют предположить, что визуальная информация через зрительный нерв является ключом к контролю дисперсии пигмента, а не наличие только синусовых желез (Shibley, 1968).То есть визуальные сигналы, проходящие к зрительной доле, кажутся решающими в контроле ответов (предположительно, через другие источники гормонов, не связанные с стебельками глаз). Поэтому необходимо проверить, как визуальная информация и роль гормонов связаны у крабов и других животных, чтобы понять пути, которыми контролируются изменения. Однако у всех видов часто предполагается, но, по-видимому, никогда не проверялось, что изменение окраски ночью также может быть механизмом экономии энергии.
Однако у всех видов часто предполагается, но, по-видимому, никогда не проверялось, что изменение окраски ночью также может быть механизмом экономии энергии. , 2013, 2014a). Это может быть связано с модификацией ритмов изменения окраски, связанных с другими факторами, такими как приливы и лунные циклы, как это имеет место у некоторых видов Uca (Fingerman, 1956; Fingerman et al., 1958), но в равной степени могут отражать различия в способности некоторых людей изменять цвет в зависимости от их состояния. Другие затраты и ограничения также могут быть связаны с изменением цвета, включая конкурирующие требования, такие как камуфляж, терморегуляция и защита от ультрафиолета.
, 2013, 2014a). Это может быть связано с модификацией ритмов изменения окраски, связанных с другими факторами, такими как приливы и лунные циклы, как это имеет место у некоторых видов Uca (Fingerman, 1956; Fingerman et al., 1958), но в равной степени могут отражать различия в способности некоторых людей изменять цвет в зависимости от их состояния. Другие затраты и ограничения также могут быть связаны с изменением цвета, включая конкурирующие требования, такие как камуфляж, терморегуляция и защита от ультрафиолета. В некотором отношении это отсутствие доказательств неудивительно. Самая последняя работа по изучению эффективности маскировки проводилась в искусственных системах с искусственными раздражителями (например, Bond and Kamil, 2002; Cuthill et al., 2005; Merilaita and Lind, 2005; Webster et al., 2013) и даже в В дикой природе только недавно исследования показали, что уровень маскировки животного напрямую влияет на вероятность нападения (Troscianko et al., 2016). Тем не менее, необходимы контролируемые эксперименты по проверке того, как изменение цвета влияет на риск нападения хищников.
В некотором отношении это отсутствие доказательств неудивительно. Самая последняя работа по изучению эффективности маскировки проводилась в искусственных системах с искусственными раздражителями (например, Bond and Kamil, 2002; Cuthill et al., 2005; Merilaita and Lind, 2005; Webster et al., 2013) и даже в В дикой природе только недавно исследования показали, что уровень маскировки животного напрямую влияет на вероятность нападения (Troscianko et al., 2016). Тем не менее, необходимы контролируемые эксперименты по проверке того, как изменение цвета влияет на риск нападения хищников. Потенциально, если спектр искусственного света сильно отличается от дневного света, то животные, такие как крабы, могут менять цвет ночью в неестественно ярких условиях на внешний вид, который не соответствует фону при естественном освещении в течение дня, а это означает, что изменение цвета ночью фактически уменьшается. дневной камуфляж.Еще одним соображением является температура и кислотность океана, связанные с изменением климата. Имеются данные о том, что изменение цвета крабов может зависеть от температуры (например, Powell, 1962b), а недавняя работа показала, как подкисление океана может влиять на поведенческие и физиологические реакции личинок рыб (Munday et al., 2009). Как изменения климата и океанов повлияют на способность животных менять цвет, еще неизвестно.
Потенциально, если спектр искусственного света сильно отличается от дневного света, то животные, такие как крабы, могут менять цвет ночью в неестественно ярких условиях на внешний вид, который не соответствует фону при естественном освещении в течение дня, а это означает, что изменение цвета ночью фактически уменьшается. дневной камуфляж.Еще одним соображением является температура и кислотность океана, связанные с изменением климата. Имеются данные о том, что изменение цвета крабов может зависеть от температуры (например, Powell, 1962b), а недавняя работа показала, как подкисление океана может влиять на поведенческие и физиологические реакции личинок рыб (Munday et al., 2009). Как изменения климата и океанов повлияют на способность животных менять цвет, еще неизвестно. В конечном счете, этот предмет должен оставаться прекрасным примером того, как исследования могут сочетать ряд междисциплинарных методов, от клеточных исследований до моделирования зрения и поведенческих экспериментов, и решать проблемы, начиная от эволюционно-экологических вопросов «сверху вниз» и заканчивая «сверху вниз». механистические вопросы «снизу вверх» (Kemp et al., 2015).
В конечном счете, этот предмет должен оставаться прекрасным примером того, как исследования могут сочетать ряд междисциплинарных методов, от клеточных исследований до моделирования зрения и поведенческих экспериментов, и решать проблемы, начиная от эволюционно-экологических вопросов «сверху вниз» и заканчивая «сверху вниз». механистические вопросы «снизу вверх» (Kemp et al., 2015).
 О ночном изменении окраски краба-горошки ( Pinnotheres veterum ). Природа 117, 415–416. дои: 10.1038/117415b0
О ночном изменении окраски краба-горошки ( Pinnotheres veterum ). Природа 117, 415–416. дои: 10.1038/117415b0 doi: 10.1086/physzool.21.4.30152016
doi: 10.1086/physzool.21.4.30152016 дои: 10.1117/12.828492
дои: 10.1117/12.828492 , Хемми Дж. М. и Бэквелл П. Р. Ю. (2008).Изменение окраски и цвета краба-скрипача, Uca capricornis : описательное исследование. PLoS ONE 3:e1629. doi: 10.1371/journal.pone.0001629
, Хемми Дж. М. и Бэквелл П. Р. Ю. (2008).Изменение окраски и цвета краба-скрипача, Uca capricornis : описательное исследование. PLoS ONE 3:e1629. doi: 10.1371/journal.pone.0001629 (1973). Поведение хроматофоров краба-скрипача Uca pugilator и карликового рака Cambarellus shufeldti в ответ на синтетический Pandalus красный пигмент-концентрирующий гормон. Ген. комп. Эндокрин. 20, 589–592. дои: 10.1016/0016-6480(73)
(1973). Поведение хроматофоров краба-скрипача Uca pugilator и карликового рака Cambarellus shufeldti в ответ на синтетический Pandalus красный пигмент-концентрирующий гормон. Ген. комп. Эндокрин. 20, 589–592. дои: 10.1016/0016-6480(73) , Трапанезе М., Маселли В., Риппа Д., Итри Ф., Аваллоне Б. и Райя П. (2014). Видение сквозь кожу: светочувствительность кожи обеспечивает криптизм у мавританского геккона. Дж. Зул. 294, 122–128. дои: 10.1111/jzo.12159
, Трапанезе М., Маселли В., Риппа Д., Итри Ф., Аваллоне Б. и Райя П. (2014). Видение сквозь кожу: светочувствительность кожи обеспечивает криптизм у мавританского геккона. Дж. Зул. 294, 122–128. дои: 10.1111/jzo.12159 T.-C., Chiao, C., Mäthger, L.M., Barbosa, A., Buresch, K.C., и Chubb, C. (2009). Динамический камуфляж головоногих: преодоление континуума между совпадением фона и разрушительной окраской. Фил. Транс. Р. Соц. В 364, 429–437. doi: 10.1098/rstb.2008.0270
T.-C., Chiao, C., Mäthger, L.M., Barbosa, A., Buresch, K.C., и Chubb, C. (2009). Динамический камуфляж головоногих: преодоление континуума между совпадением фона и разрушительной окраской. Фил. Транс. Р. Соц. В 364, 429–437. doi: 10.1098/rstb.2008.0270 Курс. Зоол. 61, 739–748. doi: 10.1093/czoolo/61.4.739
Курс. Зоол. 61, 739–748. doi: 10.1093/czoolo/61.4.739 Дж., Герберштейн, М. Е., Флейшман, Л. Дж., Эндлер, Дж. А., Беннетт, А. Т. Д., и Уайтинг, М. Дж. (2015). Интегративная основа для оценки окраски в природе. утра. Нац. 185, 705–724. дои: 10.1086/681021
Дж., Герберштейн, М. Е., Флейшман, Л. Дж., Эндлер, Дж. А., Беннетт, А. Т. Д., и Уайтинг, М. Дж. (2015). Интегративная основа для оценки окраски в природе. утра. Нац. 185, 705–724. дои: 10.1086/681021 2010.0223
2010.0223 В., Хекстра Х.Е. и Д’Агостино С.Л. (2003). Генетическая основа адаптивного меланизма у карманных мышей. Проц. Натл. акад. науч.США 100, 5268–5273. doi: 10.1073/pnas.0431157100
В., Хекстра Х.Е. и Д’Агостино С.Л. (2003). Генетическая основа адаптивного меланизма у карманных мышей. Проц. Натл. акад. науч.США 100, 5268–5273. doi: 10.1073/pnas.0431157100 doi: 10.1890/0012-9658(2001)082[2961:DVCIJM]2.0.CO;2
doi: 10.1890/0012-9658(2001)082[2961:DVCIJM]2.0.CO;2 PLoS ONE 10:e0136260. doi: 10.1371/journal.pone.0136260
PLoS ONE 10:e0136260. doi: 10.1371/journal.pone.0136260 , Пей Ронг, К., и Тодд, П. А. (2013). Изменение цвета и маскировка у рогатого краба-призрака Ocypode ceratophthalmus . Биол. Дж. Линн. соц. 109, 257–270. doi: 10.1111/bij.12039
, Пей Ронг, К., и Тодд, П. А. (2013). Изменение цвета и маскировка у рогатого краба-призрака Ocypode ceratophthalmus . Биол. Дж. Линн. соц. 109, 257–270. doi: 10.1111/bij.12039 doi: 10.1086/physzool.2.4.30152972
doi: 10.1086/physzool.2.4.30152972 Онтогенетические сдвиги в строении панциря и/или окраске литоральных и сублиторальных брахиурановых крабов. Зоопарк Raffles Bullet. 57, 543–550.
Онтогенетические сдвиги в строении панциря и/или окраске литоральных и сублиторальных брахиурановых крабов. Зоопарк Raffles Bullet. 57, 543–550. Проц. Р. Соц. В 279, 2386–2390. doi: 10.1098/rspb.2012.0026
Проц. Р. Соц. В 279, 2386–2390. doi: 10.1098/rspb.2012.0026 1098/rstb.2008.0264
1098/rstb.2008.0264 Фенотип организма будет включать не только наблюдаемые особенности, такие как морфология, но также будет включать молекулы и структуры, такие как РНК и белки, продуцируемые генами; это называется «молекулярным фенотипом».
Фенотип организма будет включать не только наблюдаемые особенности, такие как морфология, но также будет включать молекулы и структуры, такие как РНК и белки, продуцируемые генами; это называется «молекулярным фенотипом». Ген b является рецессивным признаком. Тестовое скрещивание двух растений, гетерозиготных (Bb) по признаку пурпурной окраски лепестков, даст 3 потомства с признаком пурпурных цветков (BB и Bb) и 1 потомство с признаком белых цветков ( бб). В этом примере соотношение фенотипов составляет 3:1.
Ген b является рецессивным признаком. Тестовое скрещивание двух растений, гетерозиготных (Bb) по признаку пурпурной окраски лепестков, даст 3 потомства с признаком пурпурных цветков (BB и Bb) и 1 потомство с признаком белых цветков ( бб). В этом примере соотношение фенотипов составляет 3:1. Генотип определяется как набор генов, экспрессия которых определяет характеристику или признак организма. Проще говоря, генотип — это генетический вклад в фенотип.
Генотип определяется как набор генов, экспрессия которых определяет характеристику или признак организма. Проще говоря, генотип — это генетический вклад в фенотип. о., гомозиготный рецессивный. Генотип организма является основной детерминантой фенотипа организма.
о., гомозиготный рецессивный. Генотип организма является основной детерминантой фенотипа организма.
 Когда пара аллелей, определяющих тот или иной признак, состоит из одних и тех же генов, т.е. AA или аа, признак описан как гомозиготный. Когда аллельный состав состоит из разных генов, т.е. Аа, признак описан как гетерозиготный. Наличие доминантного аллеля, т.е. АА или Аа, приведет к проявлению признака ( А ), тогда как отсутствие доминантного аллеля, т.е.е. а.о., приведут к выражению другого признака ( a ). Это случай полного доминирования, и он соответствует менделевскому наследованию.
Когда пара аллелей, определяющих тот или иной признак, состоит из одних и тех же генов, т.е. AA или аа, признак описан как гомозиготный. Когда аллельный состав состоит из разных генов, т.е. Аа, признак описан как гетерозиготный. Наличие доминантного аллеля, т.е. АА или Аа, приведет к проявлению признака ( А ), тогда как отсутствие доминантного аллеля, т.е.е. а.о., приведут к выражению другого признака ( a ). Это случай полного доминирования, и он соответствует менделевскому наследованию. Это связано с тем, что доминантный аллель будет выражен лишь частично. В результате гетерозиготное потомство будет демонстрировать фенотип, промежуточный по отношению к фенотипу родителей.
Это связано с тем, что доминантный аллель будет выражен лишь частично. В результате гетерозиготное потомство будет демонстрировать фенотип, промежуточный по отношению к фенотипу родителей. Его трансгрессивный фенотип может быть полезным или вредным в зависимости от того, как он влияет на общую приспособленность потомства. Формирование крайних фенотипов называется трансгрессивной сегрегацией . Примером гибрида с экстремальным фенотипом является потомство от скрещивания Helianthus annuus и Helianthus petiolaris . Два вида подсолнечника дали гибриды с трансгрессивными признаками. Гибриды, в отличие от их родителей, способны процветать в областях, где их родители не могут.Они способны выживать в песчаных дюнах и солончаках. (Ref.1)
Его трансгрессивный фенотип может быть полезным или вредным в зависимости от того, как он влияет на общую приспособленность потомства. Формирование крайних фенотипов называется трансгрессивной сегрегацией . Примером гибрида с экстремальным фенотипом является потомство от скрещивания Helianthus annuus и Helianthus petiolaris . Два вида подсолнечника дали гибриды с трансгрессивными признаками. Гибриды, в отличие от их родителей, способны процветать в областях, где их родители не могут.Они способны выживать в песчаных дюнах и солончаках. (Ref.1)
 ), Aabb (синяя муха с рудиментарными крыльями) и aabb (черная муха с рудиментарными крыльями).
), Aabb (синяя муха с рудиментарными крыльями) и aabb (черная муха с рудиментарными крыльями). youtube.com/embed/kLpr6t4-eLI?autoplay=1″ src=»data:text/html;https://www.youtube.com/embed/kLpr6t4-eLI?autoplay=1;base64,PGJvZHkgc3R5bGU9J3dpZHRoOjEwMCU7aGVpZ2h0OjEwMCU7bWFyZ2luOjA7cGFkZGluZzowO2JhY2tncm91bmQ6dXJsKGh0dHBzOi8vaW1nLnlvdXR1YmUuY29tL3ZpL2tMcHI2dDQtZUxJLzAuanBnKSBjZW50ZXIvMTAwJSBuby1yZXBlYXQnPjxzdHlsZT5ib2R5ey0tYnRuQmFja2dyb3VuZDpyZ2JhKDAsMCwwLC42NSk7fWJvZHk6aG92ZXJ7LS1idG5CYWNrZ3JvdW5kOnJnYmEoMCwwLDApO2N1cnNvcjpwb2ludGVyO30jcGxheUJ0bntkaXNwbGF5OmZsZXg7YWxpZ24taXRlbXM6Y2VudGVyO2p1c3RpZnktY29udGVudDpjZW50ZXI7Y2xlYXI6Ym90aDt3aWR0aDoxMDBweDtoZWlnaHQ6NzBweDtsaW5lLWhlaWdodDo3MHB4O2ZvbnQtc2l6ZTo0NXB4O2JhY2tncm91bmQ6dmFyKC0tYnRuQmFja2dyb3VuZCk7dGV4dC1hbGlnbjpjZW50ZXI7Y29sb3I6I2ZmZjtib3JkZXItcmFkaXVzOjE4cHg7dmVydGljYWwtYWxpZ246bWlkZGxlO3Bvc2l0aW9uOmFic29sdXRlO3RvcDo1MCU7bGVmdDo1MCU7bWFyZ2luLWxlZnQ6LTUwcHg7bWFyZ2luLXRvcDotMzVweH0jcGxheUFycm93e3dpZHRoOjA7aGVpZ2h0OjA7Ym9yZGVyLXRvcDoxNXB4IHNvbGlkIHRyYW5zcGFyZW50O2JvcmRlci1ib3R0b206MTVweCBzb2xpZCB0cmFuc3BhcmVudDtib3JkZXItbGVmdDoyNXB4IHNvbGlkICNmZmY7fTwvc3R5bGU+PGRpdiBpZD0ncGxheUJ0bic+PGRpdiBpZD0ncGxheUFycm93Jz48L2Rpdj48L2Rpdj48c2NyaXB0PmRvY3VtZW50LmJvZHkuYWRkRXZlbnRMaXN0ZW5lcignY2xpY2snLCBmdW5jdGlvbigpe3dpbmRvdy5wYXJlbnQucG9zdE1lc3NhZ2Uoe2FjdGlvbjogJ3BsYXlCdG5DbGlja2VkJ30sICcqJyk7fSk7PC9zY3JpcHQ+PC9ib2R5Pg==»>
youtube.com/embed/kLpr6t4-eLI?autoplay=1″ src=»data:text/html;https://www.youtube.com/embed/kLpr6t4-eLI?autoplay=1;base64,PGJvZHkgc3R5bGU9J3dpZHRoOjEwMCU7aGVpZ2h0OjEwMCU7bWFyZ2luOjA7cGFkZGluZzowO2JhY2tncm91bmQ6dXJsKGh0dHBzOi8vaW1nLnlvdXR1YmUuY29tL3ZpL2tMcHI2dDQtZUxJLzAuanBnKSBjZW50ZXIvMTAwJSBuby1yZXBlYXQnPjxzdHlsZT5ib2R5ey0tYnRuQmFja2dyb3VuZDpyZ2JhKDAsMCwwLC42NSk7fWJvZHk6aG92ZXJ7LS1idG5CYWNrZ3JvdW5kOnJnYmEoMCwwLDApO2N1cnNvcjpwb2ludGVyO30jcGxheUJ0bntkaXNwbGF5OmZsZXg7YWxpZ24taXRlbXM6Y2VudGVyO2p1c3RpZnktY29udGVudDpjZW50ZXI7Y2xlYXI6Ym90aDt3aWR0aDoxMDBweDtoZWlnaHQ6NzBweDtsaW5lLWhlaWdodDo3MHB4O2ZvbnQtc2l6ZTo0NXB4O2JhY2tncm91bmQ6dmFyKC0tYnRuQmFja2dyb3VuZCk7dGV4dC1hbGlnbjpjZW50ZXI7Y29sb3I6I2ZmZjtib3JkZXItcmFkaXVzOjE4cHg7dmVydGljYWwtYWxpZ246bWlkZGxlO3Bvc2l0aW9uOmFic29sdXRlO3RvcDo1MCU7bGVmdDo1MCU7bWFyZ2luLWxlZnQ6LTUwcHg7bWFyZ2luLXRvcDotMzVweH0jcGxheUFycm93e3dpZHRoOjA7aGVpZ2h0OjA7Ym9yZGVyLXRvcDoxNXB4IHNvbGlkIHRyYW5zcGFyZW50O2JvcmRlci1ib3R0b206MTVweCBzb2xpZCB0cmFuc3BhcmVudDtib3JkZXItbGVmdDoyNXB4IHNvbGlkICNmZmY7fTwvc3R5bGU+PGRpdiBpZD0ncGxheUJ0bic+PGRpdiBpZD0ncGxheUFycm93Jz48L2Rpdj48L2Rpdj48c2NyaXB0PmRvY3VtZW50LmJvZHkuYWRkRXZlbnRMaXN0ZW5lcignY2xpY2snLCBmdW5jdGlvbigpe3dpbmRvdy5wYXJlbnQucG9zdE1lc3NhZ2Uoe2FjdGlvbjogJ3BsYXlCdG5DbGlja2VkJ30sICcqJyk7fSk7PC9zY3JpcHQ+PC9ib2R5Pg==»>